

Science精选丨高质量基因组探究适应性进化重复演化机制
基因组重组抑制区域(如染色体倒位)可能促进适应。这类区域能维持多个连锁且共适应基因座的复合体,从而提升适应性。近年来越来越多证据显示,适应可能涉及大尺度重组抑制基因组区域,这些区域作为大效应基因座发挥作用,但倒位与转座元件以外的结构变异在适应中的作用仍待探索。
近日,多国科学家团队合作,在国际著名顶刊《Science》发表了题为“Adaptation repeatedly uses complex structural genomic variation”的研究论文,利用单倍型组装基因组与群体测序数据,探究了竹节虫局部适应的遗传基础,重点关注了结构变异的作用及进化可重复性。

一、竹节虫生态型间的体色差异
无翅植食性竹节虫Timema cristinae表现出两种生态型,第一种生态型栖息于蔷薇科小型植物Adenostoma fasciculatum,通常表现为带有背部白色纵条纹的绿色体色(以下简称"条纹型")。这种形态在Adenostoma的针状叶片上具有伪装效果。第二种生态型栖息于鼠李科大型乔木植物Ceanothus spinosus,通常表现为无条纹的纯绿色体色(以下简称"绿色型"),在Ceanothus的宽叶上形成伪装。
二、基因组组装与两地生态型的种群历史
作者通过结合PacBio测序和Omni-C测序技术,为T. cristinae的四个个体(Refugio和Hwy154两座山脉各取条纹型和绿色型各一例)完成了单倍型分型的染色体级别基因组组装(共获得八套独立组装数据)。
作者首先将GBS数据比对至Refugio条纹型基因组单倍型。基于获得的204,442个SNP位点进行群体基因组分析,结果证实了两地间极低的基因流(每世代<1个迁移个体)。估算Refugio与Hwy154种群约在123万年前分化,印证了两地种群在相当长时间内基本独立进化。
图1:Refugio和Hwy154两座山脉条纹型和绿色型竹节虫
三、Refugio地区体色模式与大型倒位易位相关
作者利用新组装的Refugio参考基因组和218只T. cristinae竹节虫(98只绿色型,120只条纹型)的GBS数据,分析了Refugio种群内体色模式的遗传基础。GWA分析显示,一个约43Mbp的区域关联信号集中在chr8上。区域两端各有一个显著关联峰,形成"悬索桥"结构,这种结构通常出现在染色体倒位或其他抑制重组的结构变异中。通过比较Refugio单倍型基因组组装,作者发现该区域并非简单倒位,而是一个约9-Mbp片段的染色体内易位——该片段发生倒位后移位约34 Mbp,恰好与GWA信号重合。因此,Refugio种群体色变异由倒位与易位共同构成的复杂结构变异驱动,其中易位起关键作用。
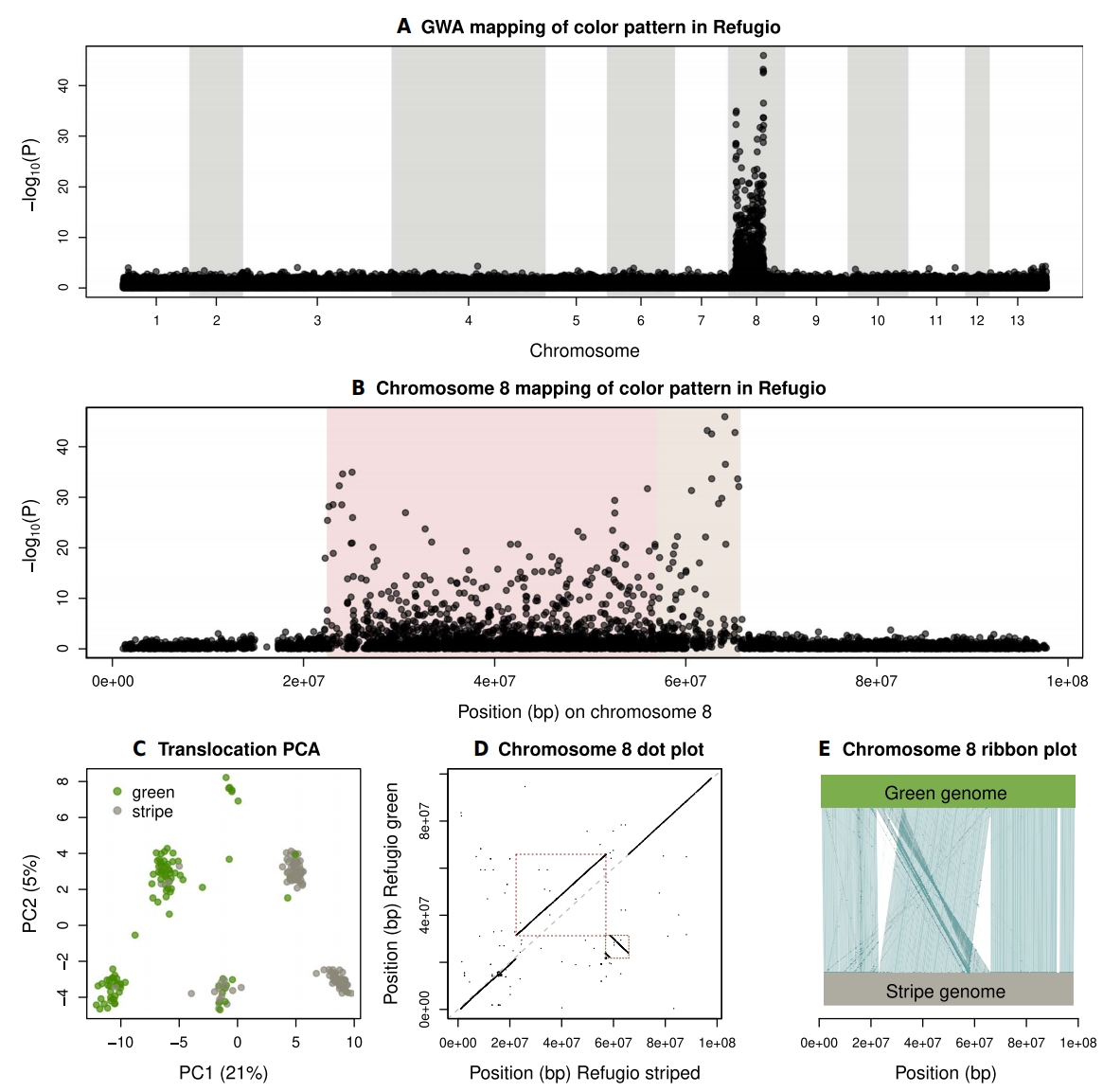
图2:Refugio山脉竹节虫颜色变异的遗传学基础
四、Hwy154体色模式涉及不同的倒位易位
先前对Hwy154的竹节虫研究也发现GWA信号集中在chr8上,但其在染色体中的具体位置与上文Refugio结果有所不同。作者对Hwy154的538份GBS数据重复了与Refugio相同的GWA分析步骤,首次利用单倍型参考基因组定位了该山脉体色模式的遗传基础。
作者在Hwy154的chr8上发现跨越约80 Mbp的多个关联峰。鉴定出一个约15 Mbp的结构变异,该变异与两个体色模式的GWA峰相吻合。比较组装结果显示,用于单倍型基因组组装的Hwy154条纹型竹节虫在该结构变异区域呈杂合状态,所以作者重点分析携带该变异的单倍型。
这一复杂SV包含一个约7 Mbp的区域,该区域发生倒位并易位至同一染色体的相邻区域约8 Mbp处,同时在约7 Mbp区域内还存在一个较小的次级易位。因此再次发现了复杂重排和易位的证据,但这里涉及的是约15 Mbp区域,与Refugio约43 Mbp的大区域形成对比。该基因组区域与之前基于非单倍型基因组研究中发现的Mel-Stripe2基因座大致重合。
这些综合结果表明,条纹型和绿色型形态的进化反复利用了复杂结构变异,尽管不同山脉的变异起源不同。
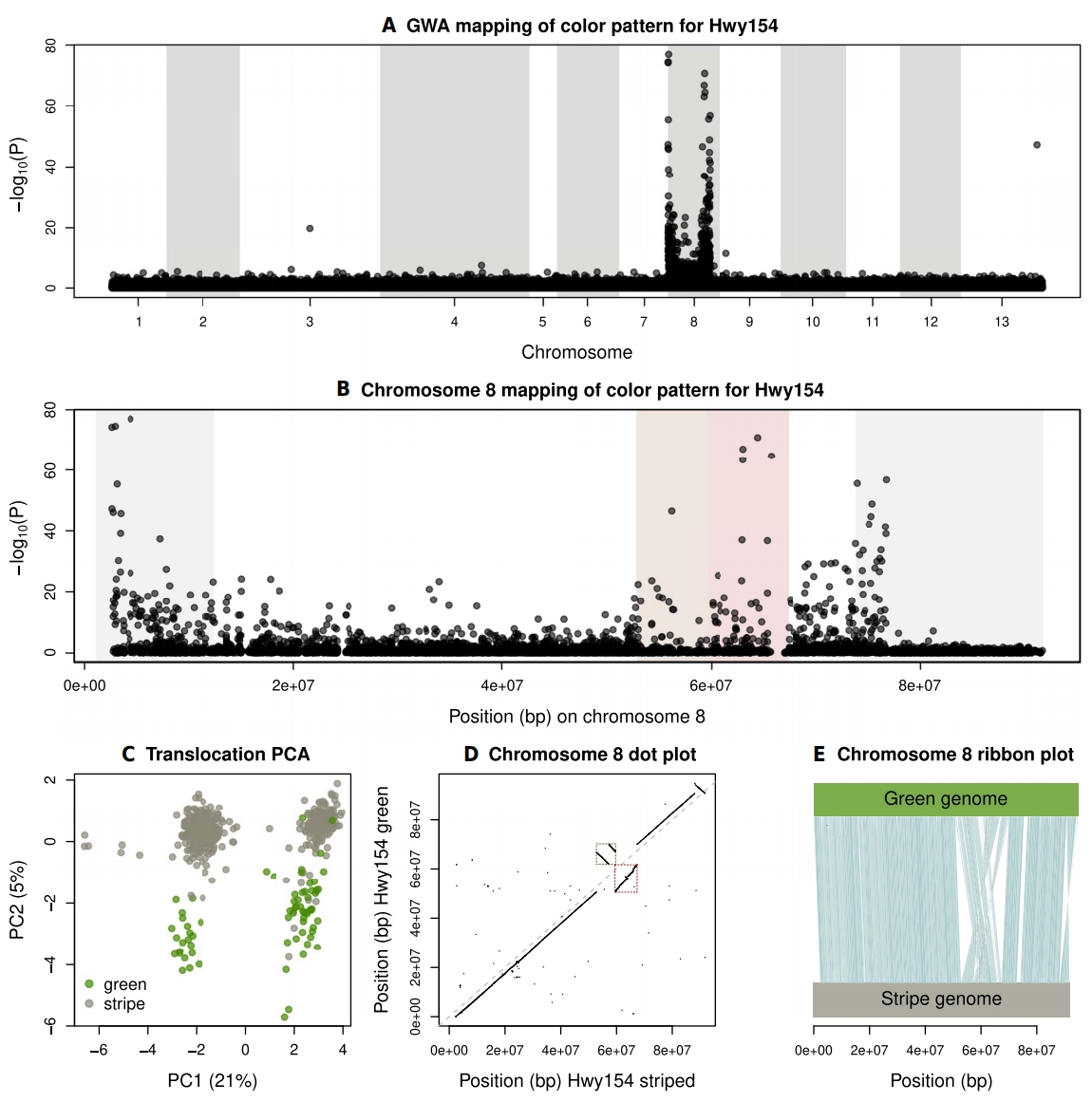
图3:Hwy154山脉竹节虫颜色变异的遗传学基础
五、结构变异的遗传与功能解析
基于Refugio和Hwy154两地绿色型与条纹型基因组的同源基因分析表明,两座山脉的倒位易位事件既存在差异又部分重叠。Refugio的9 Mbp倒位区与34 Mbp共线区的小片段,与Hwy154易位区的8 Mbp共线区存在重叠,而Refugio易位区的主体共线部分超出Hwy154易位边界,但Hwy154的7 Mbp倒位区与Refugio易位无交集,表明两地倒位易位应该为独立起源。
Refugio和Hwy154易位倒位区功能基因分析研究显示,直翅目体色调控关键基因(corazonin基因)在Refugio定位在断点外侧,而Hwy154则位于倒位区内,另外调控体色黑化的蜕皮激素激酶基因在两地个体中均存在双拷贝,转座元件分布显示断点区检测到hAT等TE元件。
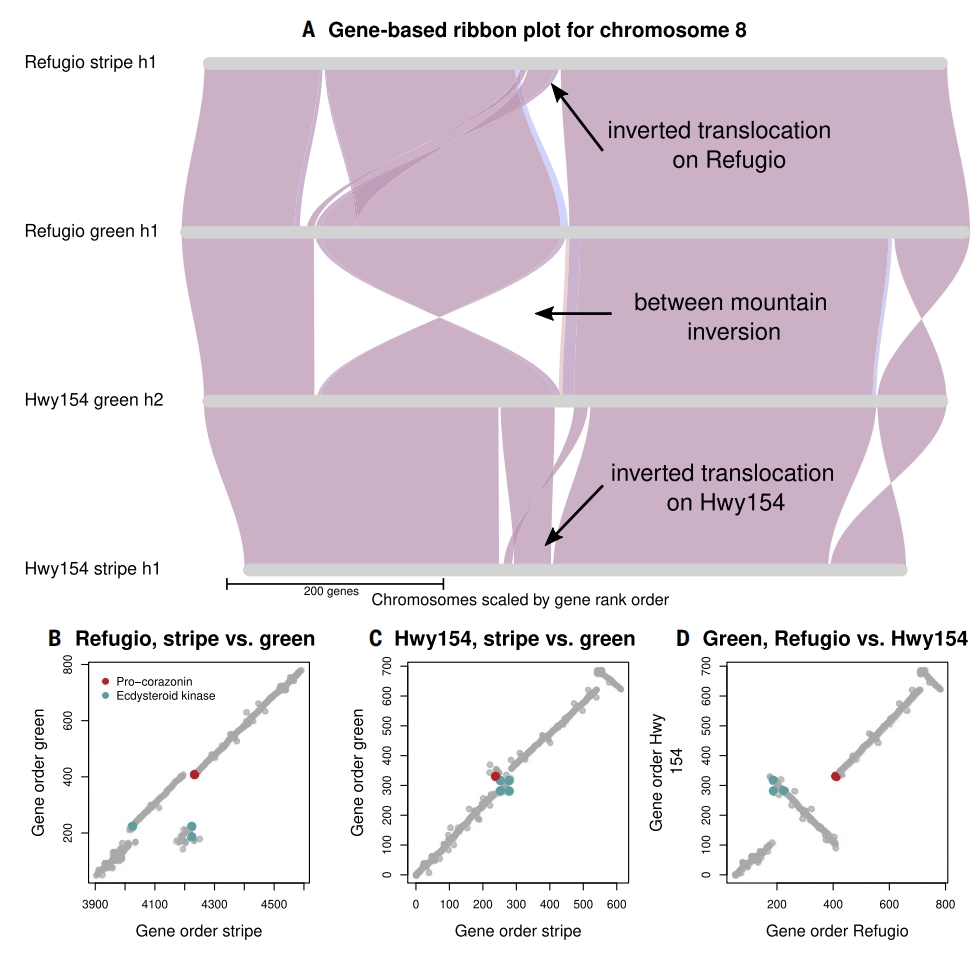
图4:Refugio与Hwy154种群体色模式结构变异的比较基因组学分析
六、自然选择作用验证
之前的相关研究均推断,这些结构变异因与体色模式的关联而受到自然选择的作用。为验证这一假说,作者利用Refugio地区形态频率的自然渐变分布,分析基于218只用于全基因组关联分析(GWA)的T. cristinae个体,估算易位等位基因频率的地理渐变宽度为576米。该渐变曲线呈S型,尾部无明显的阶梯或指数衰减,此模式符合选择-扩散平衡模型。通过杂交带中心的LD估算扩散距离,显示除易位外的全基因组位点均未呈现渐变变异,符合中性进化预期。易位与其他位点的鲜明对比,进一步佐证了控制体色模式的易位受到自然选择的结论。
在确定Refugio地区易位受到自然选择作用后,作者进一步研究其进化历史,检验其是否可能通过基因渗入(introgression)从其他近缘物种传入T. cristinae。作者利用TreeMix和F3统计量分析了T. cristinae与北加州其他Timema物种间的基因渗入信号,分析结果显示无渗入证据。所以推断Refugio地区隐蔽色的本地适应涉及一个受渐变分布上分歧选择作用的易位。Hwy154的隐蔽色则由另一独立易位控制,同样未检测到近缘物种渗入信号。
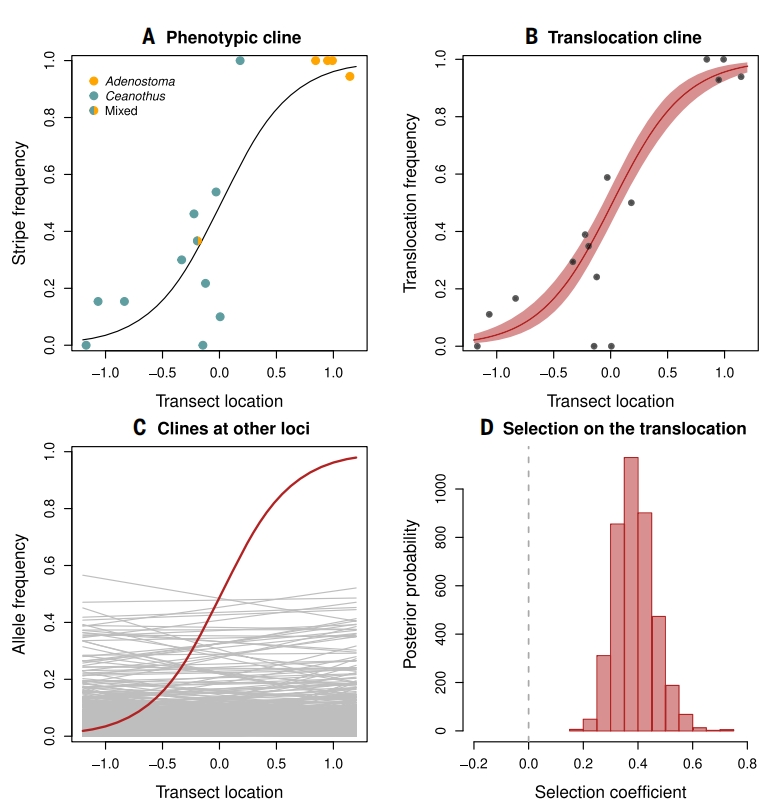
图5:Refugio种群渐变分布上易位受自然选择的证据
七、全基因组结构变异的可检测性差异
作者进一步扩展研究,检验“结构变异普遍难以通过短读长测序数据进行群体遗传学检测”的假说。这一假说认为:单倍型基因组组装对比可显著提升检测灵敏度,甚至能揭示单个个体中的分离结构变异。
为验证此观点,作者首先对Refugio和Hwy154的单倍型参考基因组进行了全基因组比对(chr8以外区域),鉴定出多染色体结构变异。Refugio种群中存在7个大型染色体倒位,针对这些倒位区域的PCA分析显示,其中3个倒位存在遗传聚类,而另4个则无此信号。这表明约半数倒位若仅依赖群体遗传学数据将难以检测。Hwy154种群中6个大型染色体重排也呈现类似模式,部分倒位或易位倒位无遗传聚类信号。这种检测困难可能源于倒位近期起源、低频基因交换破坏分化信号,或二者共同作用。
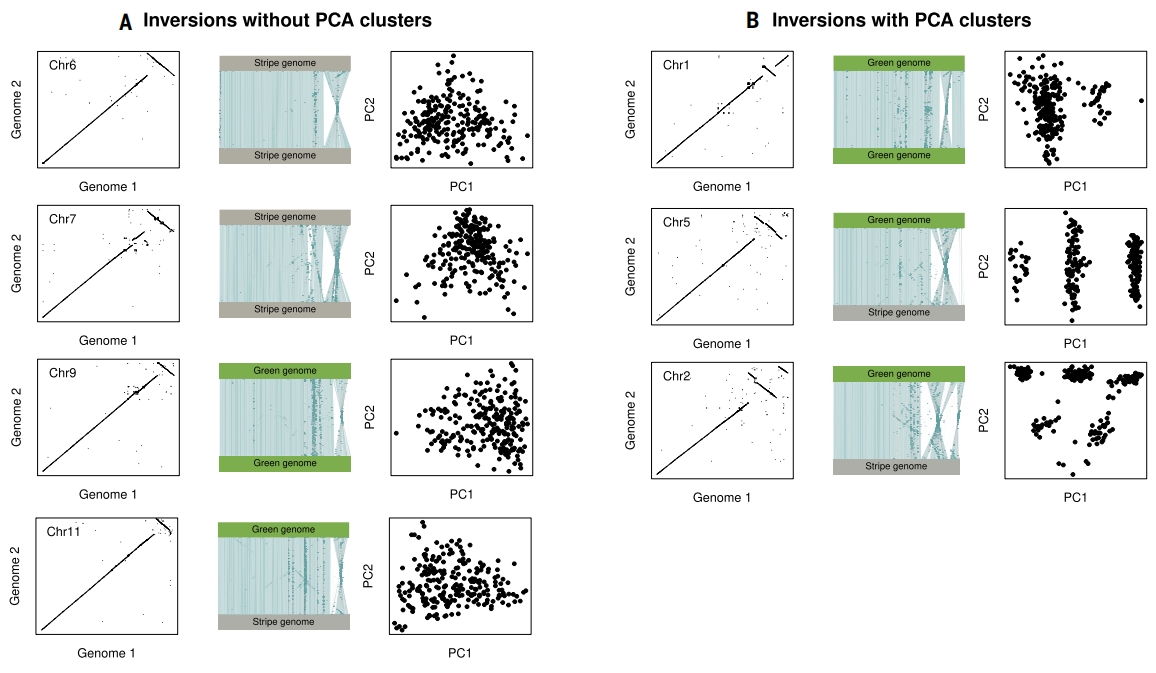
图6:Refugio地区T. cristinae全基因组大型倒位的群体遗传模式总结
结语
本研究揭示了竹节虫适应性多态性背后的结构变异,其由自然选择与基因流的平衡维持,本研究发现同一物种不同种群的适应性分化可能由不同类型和复杂程度的结构变异反复驱动。本研究的发现表明,结构变异可能是一种丰富多样且广泛存在的遗传变异来源,为进化提供持续动力。更重要的是,这类变异的反复出现可能为适应性进化提供了某种程度的可预测性。随着基因组学技术的持续进步,这一假说将能在更多生物类群和进化情境中得到验证。
华命生物产品服务一览

华命生物目前已开通微信公众号、抖音、知乎、B站、小红书等线上平台,欢迎感兴趣的老师扫码关注了解更多内容!
