

NC重磅丨刘建全团队解析古异源多倍化驱动杨属与柳属的辐射演化
多倍化(WGD)在被子植物中普遍存在,并对其演化起着关键作用。多倍体可通过异源多倍化或同源多倍化形成,由于染色体的大规模重排和亲本谱系的可能灭绝,识别经历过再二倍化的古老异源多倍化事件并区分其亚基因组具有挑战性。再二倍化的异源多倍体需要基于基因含量的残留差异、表达偏好和重复DNA特征来推断亚基因组。但当古老异源多倍体呈现高度同质化的亚基因组时,这些方法便难以奏效。因此对于古老异源多倍体亚基因组的对比性演化及其在高等分类类群中创新性状形成、物种多样化及明显生态位分化等方面的贡献,目前仍缺乏深入研究。
近日,四川大学/兰州大学刘建全教授和四川大学马涛教授团队,在国际著名期刊《Nature Communications》上,发表了标题为“Ancient allopolyploidy and specific subgenomic evolution drove the radiation of poplars and willows”的研究论文,作者以木本模式类群杨柳科(Salicaceae)为对象,构建涵盖其全部三个亚科的基因组,发现杨柳科存在一个共享的古老异源多倍化事件,随后发生分化的核型与亚基因组进化,这种特异性进化推动了物种丰富的杨属和柳属创新性状的起源。

一、杨柳科物种基因组组装和WGD事件解析
作者整合了Nanopore测序数据、NGS测序数据以及Hi-C数据,成功组装了杨柳科八个属种的染色体级别基因组,包括柳亚科(Salicoideae)的七个物种(Dovyalis caffra、Scolopia chinensis、Xylosma longifolia、Flacourtia jangomas、Itoa orientalis、Idesia polycarpa和Salix rehderiana)以及天料木亚科(Samydoideae)的一个物种(Casearia decandra)。另外利用PacBio HiFi测序数据和Hi-C数据,还完成了鞘冠木亚科(Scyphostegioideae)物种Dianyuea turbinata的染色体级别组装。这些新组装基因组大小差异显著,其中D. turbinata(4,183 Mb;n = 18 + 1B)和S. chinensis(274 Mb;n = 11)分别具有最大和最小的基因组。Hi-C热图沿主对角线显示强烈互作信号,表明组装具有高连续性和准确性.
基于BUSCO和OrthoFinder方法鉴定了19个物种的267个单拷贝基因,作者重建了高分辨率系统发育树。该树证实杨柳科三个亚科均为单系群。在柳亚科(Salicoideae)内,作者识别出两个进化枝:一个包含染色体数n=19-21的类群(进化枝I),另一个染色体数较少(n=9-11;进化枝II)。
通过比较这些基因组的染色体和基因数量,作者推断"杨柳类"WGD事件可能仅限于进化枝I物种。为验证此假设,作者评估了每个基因组中旁系同源基因的Ks。结果表明,尽管所有物种都经历了古老的WGD(γ事件),但D. turbinata和进化枝I物种还经历了近期的独立WGD事件,表明其多倍化起源独立。此外进化枝I物种内部(2:2)及与进化枝II物种间(2:1)的良好共线性关系进一步支持这一结论。这些结果一致表明"杨柳类"WGD事件发生在进化枝I物种的祖先分支上。
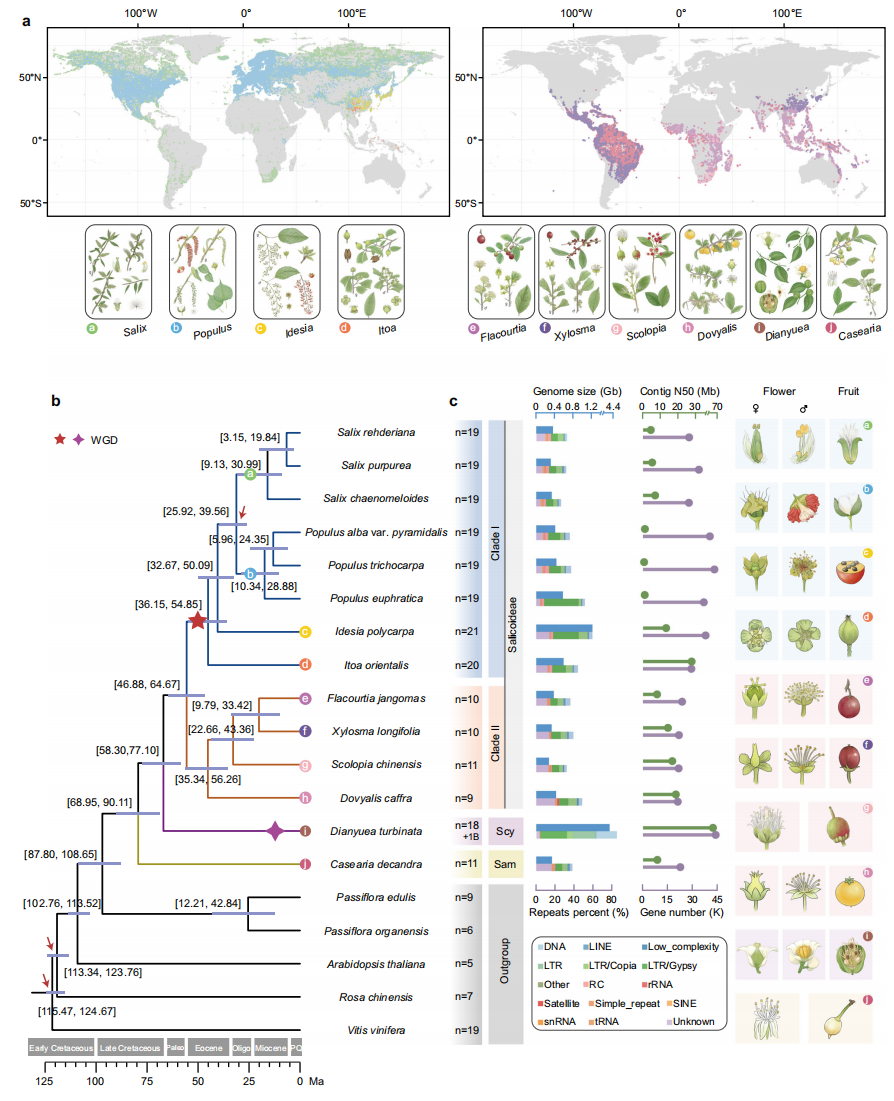
图1:杨柳科杨属、柳属及相关属的分布与系统发育关系
二、核型进化与染色体重排
利用这些物种间共享的完整染色体和/或类染色体同线性区块,作者重建了杨柳科祖先核型(ASK),其包含11条完整原型染色体,而非先前推断的染色体基数10。基于"杨柳类"WGD和基因组重排事件,作者进一步追溯了它们的进化关系,其结果与系统发育分析高度吻合。
作者进一步推断柳亚科进化枝I物种的祖先核型在"杨柳类"WGD后基染色体数为21。此外,作者还预测了多个原型染色体间的RTA事件。这些全基因组重组在I. polycarpa中完全保留且未发生其他大尺度融合/断裂事件,表明它们的共同祖先在再二倍化过程中已拥有相对稳定的核型。杨属和柳属的祖先也经历了一系列染色体重排:通过重复的原型染色体3、7和11的连续嵌套染色体融合(NCF)形成一条古老融合染色体,该染色体后续发生两次独立RTA事件,最终导致杨属和柳属形成19条染色体的核型。
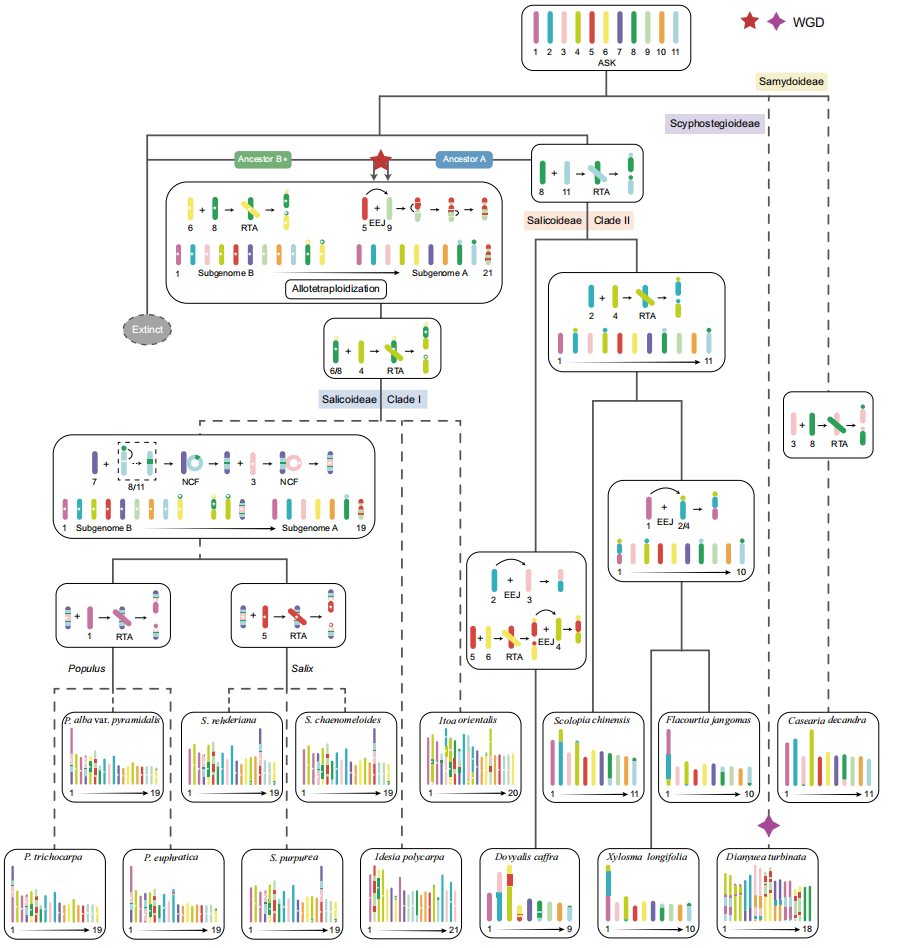
图2:杨属、柳属及相关类群从共同祖先核型出发的核型进化与系统关系
三、"杨柳类"WGD事件属于隐性异源多倍化
通过整合杨柳科物种祖先核型(ASK)与现存染色体的对应关系、系统发育分析及亚基因组划分,作者发现了有力证据表明"杨柳类"WGD是一次隐性异源多倍化事件。根据重建的11条原型染色体,作者鉴定了1545个完整同源基因群(HGGs),其中进化枝I所有物种均保留了"杨柳类"WGD产生的旁系同源基因,而其他物种仅保留直系同源基因。基于ASTRAL溯祖方法构建的系统发育树显示,进化枝II的基因与进化枝I物种"杨柳类"旁系同源基因的一个拷贝亲缘关系更近。这一关系得到所有11条原型染色体上86.7%-94.1%完整HGGs的支持。此外,作者发现进化枝II祖先中发生的原型染色体8与11的RTA事件,仅存在于进化枝I所有物种的一条染色体中,进一步支持其异源多倍化起源。
根据与进化枝II的系统发育关系,作者将进化枝I物种的基因组划分为两个亚基因组:A亚基因组包含与进化枝II亲缘关系密切的染色体,B亚基因组则由"杨柳类"WGD产生的旁系同源染色体组成。使用溯祖法和串联法分别基于A、B亚基因组重建的系统发育树均呈现高度支持的一致拓扑结构。这些结果表明进化枝I物种的"杨柳类"事件是一次异源多倍化事件,其中一个亲本是进化枝II物种的共同祖先。
为识别"杨柳类"异源多倍化的另一个可能亲本,作者进一步组装了柳亚科四个附加物种(Azara serrata、Prockia crucis、Banara guianensis和Homalium cochinchinense)的参考基因组。将这四个属的基因序列添加到HGGs中,分别构建溯祖法和串联法系统发育树。两棵树均显示这些柳亚科新属聚类在进化枝II内,而D. turbinata的两个亚基因组聚在一起,并作为进化枝I物种两个亚基因组的姐妹群,表明这些物种均非B亚基因组的直接供体。由于这些物种代表了杨柳科全部三个亚科,作者推断B亚基因组的亲本供体很可能已灭绝。而基于A、B亚基因组分化时间约66 Ma、进化枝II与A亚基因组分化约58 Ma的估算,作者推测两个亲本谱系在"杨柳类"WGD前已分化约800万年。
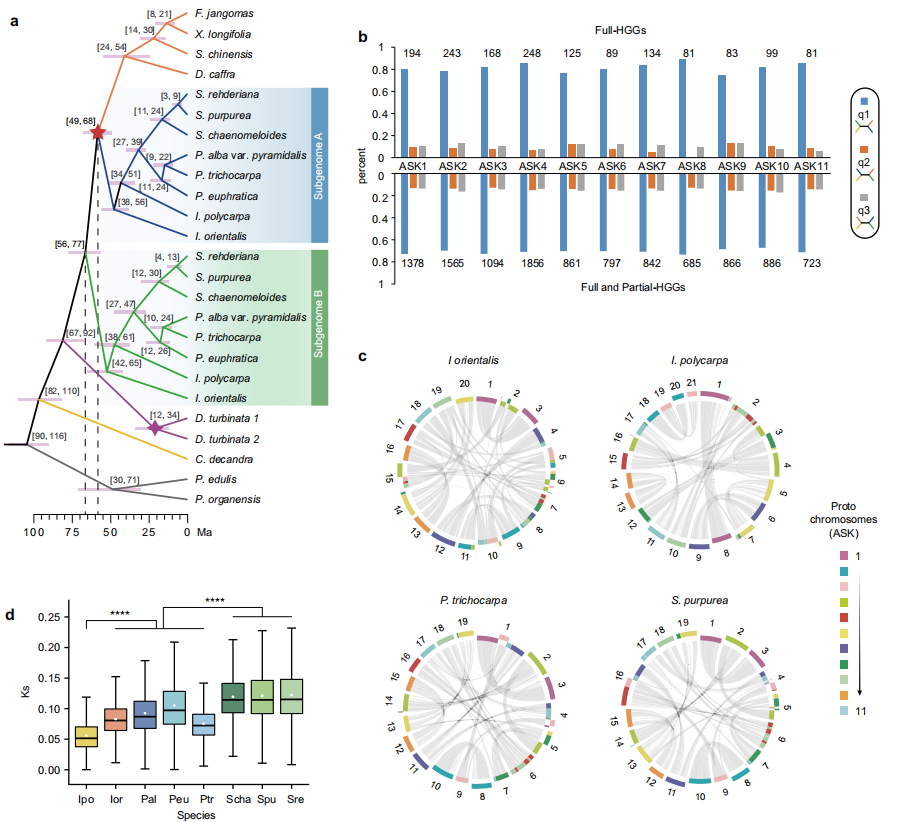
图3:杨属、柳属及其两个姐妹属异源多倍化事件的鉴定
四、亚基因组优势与谱系特异性基因保留
多倍化后的亚基因组进化通过减少基因冗余,为性状创新和适应性进化提供关键基础。作者对23个杨柳科基因组/亚基因组的分析表明,在进化枝I物种中,A亚基因组表现出相对于B亚基因组的进化优势。"杨柳类"WGD事件后,进化枝I物种的基因保留率显著降低,且A亚基因组的保留率始终高于B亚基因组。而在表达水平方面,所有进化枝I物种中,B亚基因组的基因表达水平均低于A亚基因组。这可能与B亚基因组基因周围更高的重复序列含量有关。此外表观遗传特征方面,A亚基因组基因具有显著更高的染色质可及性和略低的甲基化水平。
作者进一步分析了进化枝I三个关键节点的基因保留模式,在全进化枝I中鉴定1989对WGD衍生基因对,这些基因对功能主要关联"光周期开花调控"、"花序发育"、"种子发育调控"、"毛状体形态发生"、"冷驯化"和"防御响应"等,这些基因可能通过适应北方多变光周期与温度促进进化枝I与祖先谱系分化。而落叶谱系中特异性保留402对基因,主要包含光周期开花相关基因。另外杨属-柳属物种中鉴定129对特有保留基因对,功能主要涉及"花器官身份维持"、"种子毛状体分化"、"红/远红光信号通路"和"植物细胞壁纤维素生物合成调控"。
这些谱系中基因保留的动态模式表明,光周期响应、开花时间、花序结构和种子特性等方面的创新性状共同促进了进化枝I物种的多样化适应。
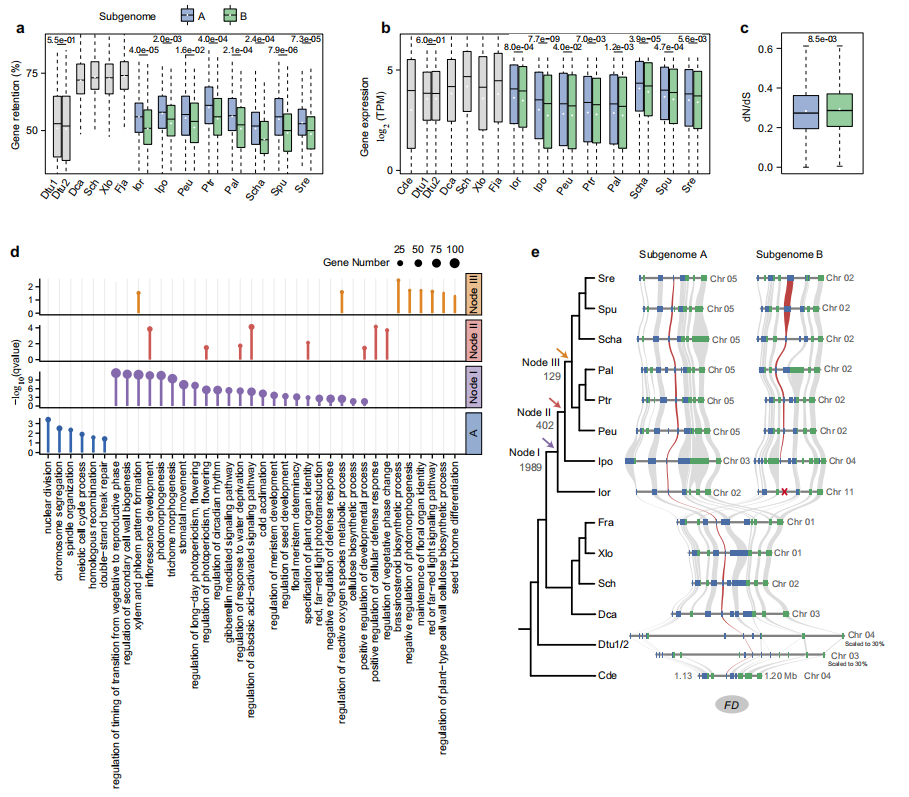
图4:杨属-柳属谱系及其两个姐妹谱系的亚基因组优势与基因保留
五、谱系特异性亚基因组表达促进杨属与柳属多性状创新
旁系同源基因的表达分化在多倍化后驱动物种辐射与进化创新中起关键作用。为探究杨柳科亚基因组表达模式与谱系分化的关联,作者聚焦于进化枝I所有物种共同保留的基因对。通过对伊桐属、山桐子属和杨属雄花、雌花及不同发育阶段果实的转录组测序与聚类分析,发现同一物种内A、B亚基因组基因对的表达偏好在不同组织和发育阶段高度一致,但物种间存在显著差异。
通过分析伊桐和山桐子从晚秋至早春叶片和芽的表达模式,作者发现:在叶片中,伊桐冬季同时表达FT1和FT2,而山桐子和杨属仅表达FT1。但在芽/茎尖中,杨属特异性表达FT1,伊桐冬季持续表达FT2,山桐子仅在晚冬表达FT2。这种分化与杨属从营养生长向生殖生长的物候转换一致,使其能在春季更早开花,而伊桐的共表达模式与其常绿无休眠特性相关,山桐子则关联春季回暖后生殖生长启动。
杨属特异性表达分化的基因对显著富集于"光周期开花"、"生物节律调控"、"花发育"、"种子发育调控"和"毛状体形态发生"等功能,转基因功能验证显示,在拟南芥中过表达杨属ZFP6或GIS3,转基因株系叶片毛状体密度约为野生型2倍,证实二者作为毛状体发育正调控因子的功能保守性。此外,在ZFP6和GIS3启动子区鉴定出4个杨属-柳属特异性CNEs。报告基因实验表明这些CNEs具有增强子活性,可能促进该谱系种子毛状体发育中的高效表达。
亚基因组表达的谱系特异性分化显著促进了杨属和柳属在开花物候和具毛小种子等性状的进化,这些性状可能为其向高纬度地区快速扩张和广泛辐射提供关键适应优势。
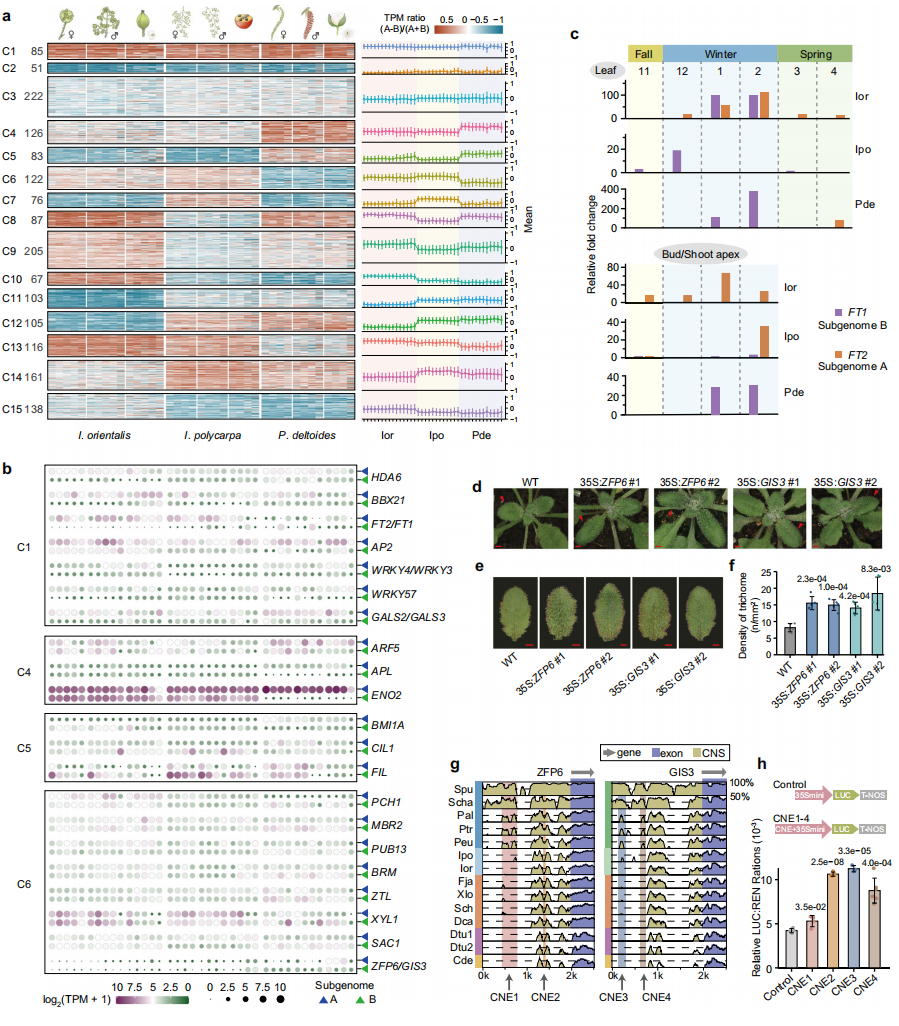
图5:杨属-柳属及其两个姐妹谱系中保留重复基因对的亚基因组表达与调控模式
结语
本研究揭示了异源多倍化后亚基因组的分化进化轨迹,这些轨迹构成了杨柳科多样化性状的基础。尽管古多倍化事件普遍存在,但对其亚基因组长期进化动态的综合分析仍十分匮乏。本研究建立的整合分析框架,结合这些类群中快速积累的基因组资源,为探究古多倍化和亚基因组进化如何塑造生命之树中的适应性辐射和生物多样性模式提供了坚实基础。最重要的是,作者发现强调需区分同源与异源多倍化事件,并关注分化的亚基因组进化,这些过程在足够长的进化时间中能够促进创新性状的出现和物种多样化。
华命生物产品服务一览

华命生物目前已开通微信公众号、抖音、知乎、B站、小红书等线上平台,欢迎感兴趣的老师扫码关注了解更多内容!
