

NP重磅 | 生姜基因组解码雌雄异熟现象
被子植物为降低自交率、增强遗传多样性,演化出多种促进异花授粉的机制,约70%的物种表现出雄蕊与雌蕊在时间上的分化,包括雄蕊先熟和雌蕊先熟两种类型。该机制通过错开花药开裂与柱头可授期的时间,有效减少自花授粉几率。以姜科山姜属Alpinia植物为例,其雄蕊先熟型与雌蕊先熟型花朵在一天内呈现出高度同步的花柱运动与花药开裂节律:清晨雌蕊先熟型接受来自雄蕊先熟型的花粉,午后则角色互换,实现双向异交。这种精密的时空协同机制不仅提升了授粉效率,也为研究植物生殖节律及其分子调控机制提供了理想的研究模型。
云南大学生物多样性研究员赵建立研究员及科研团队于2025年10月7日在著名刊物nature plants上发表了题为“Ginger genome reveals the SMPED1 gene causing sex-phase synchrony and outcrossing in a flowering plant”的研究论文,在组装出单倍型生姜基因组的基础上,进一步鉴定出关键基因 SMPED1,系统揭示了柔性花柱(flexistyly)这一特殊雌雄异熟现象的遗传与分子机制。

一、在马来良姜A. mutica 中花型的孟德尔遗传机制
作者发现,PG型(雄蕊先熟)和PA型(雌蕊先熟)花在野生种群中的比例接近1:1,表明柔性花柱受单一孟德尔遗传位点控制。为了验证该假设,作者对马来良姜和长柄山姜Alpinia kwangsiensis进行杂交,得到的F1后代中PG:PA比例为1:1;PA的自交只产生PA后代,而PG的自交则产生两种花型,PG:PA比例为3:1。柔性花柱的遗传由显性等位基因(d/D)和隐性等位基因(d/d)控制,保证了PG和PA花型在后代中的平衡比例。研究结果与其他异型雌雄异熟物种的杂交数据一致,证实了这一生殖适应机制的遗传基础。

图1:山姜属植物雌雄异熟的孟德尔遗传
二、马来良姜基因组概述
本研究构建了马来良姜的高质量单倍型解析基因组,使用PacBioHiFi测序结果进行组装,共得到两种花型的四个单倍型组装,平均长度为2.18Gb,BUSCO完整度达98.7%。每个单倍型约含5万个蛋白编码基因,呈现高度的基因共线性。基因组中可转座元件比例高达85%,主要来源于LTRs扩增,尤其是Copia元件。
三、基因组的二型花期决定区域
通过对91个个体进行重测序,研究共鉴定出1271万个SNP,基于GWAS分析,作者在第8号染色体末端发现了一个与雌雄异熟显著相关的SNP区域(约8.36kb);尽管在第13号染色体上也观察到关联信号,但由于未发现明显的连锁区域或大型转座元件插入,作者将第8号染色体确定为主要研究对象。进一步分析表明,该区域符合孟德尔遗传规律:PA型个体中参考等位基因占主导,而PG型个体则以替代等位基因为主。通过单倍型结构与重组分析,作者推算DDR区域(Duplicated Domain Region,重复结构区)的范围界定为约30kb,并在PG型中识别出一个约50kb的杂合区,该区域与PA型序列高度一致,表明其为一个半合子位点。最终作者将候选DDR区域精确界定在第8号染色体的100518121–100547025bp区间。遗传多样性分析显示,PG型DDR区域的整体多样性显著低于PA型,但作者发现其中的AmSMPED1基因在PG型中表现出更高的多样性,认为可能与LTR元件插入有关。

图2:A. mutica基因组内DDR区域的鉴定
四、雌雄异熟决定基因SMPED1 与雌雄异熟表型
研究进一步聚焦于山姜属植物中鉴定控制雌雄异熟的关键遗传区域DDR,并对其中的候选基因进行了功能探索。作者在DDR区域内锁定了两个候选基因:AmSMPED1 和AmCOR27。通过在不同光照条件(自然光与连续光)下,对开花期花药和花柱进行高精度时序转录组分析,发现只有AmSMPED1的表达节律与花药开裂和花柱运动的时序高度同步:AmSMPED1的表达变化与PG表型在早晨9点左右的花药开裂及花柱收回行为吻合;在连续光照下,PA表型的AmSMPED1表达节律则发生完全逆转,表明该基因是调控性阶段转换的核心。进一步的机制研究揭示,在PG (d/D) 表型中,AmSMPED1的显性D等位基因表达量低于隐性d,启动子活性实验证实,这是由d等位基因更高的启动子活性所致。相比之下,另一个候选基因AmCOR27以及其他DDR内基因的表达均与花部行为无关。因此,作者最终确定AmSMPED1为驱动山姜属植物雌雄异熟表型形成的关键候选基因。

图3:SMPED1决定雌雄异熟性阶段同步性的验证
五、SMPED1决定性阶段的同步性
马来良姜的花柱组织具渗透性,适合AS-ODN 处理,但花药因具次生壁而无法处理。鉴于马来良姜尚未建立转基因系统,SMPED1 基因在被子植物中高度保守,研究者以拟南芥Arabidopsis thaliana为模型探究其在花药开裂中的作用。拟南芥基因组中存在两个与 AmSMPED1 高度同源的基因,其启动子驱动 GUS 表达显示在花药中有强烈活性。作者发现,在拟南芥中构建的过表达株系花药开裂时间均较野生型提前,而双基因下调株系的花药开裂时间则显著延迟。结果表明,SMPED1促进花药开裂,进一步印证了该基因在马来良姜花药与花柱发育同步中的关键作用。
六、被子植物DDR基因的进化与SMPED1基因的保存
系统基因组学分析显示,芭蕉科是姜目中最早分化的类群,其基因组中存在DDR结构。约3250万年前,姜科植物经历了一次全基因组复制(WGD),产生了两份DDR拷贝,并在进化过程中分化为I型和II型DDR。LTR序列在AmSMPED1与AmCOR27之间的插入与缺失事件驱动了不同形态(PG型与PA型)DDR的形成,其中PG型约起源于406万年前。研究发现,各物种间DDR的基因排列高度共线,反映出其祖先基因组结构的保守性。同时,SMPED1基因广泛分布于几乎所有被子植物,其演化时间与基底被子植物的出现大致同步。尽管SMPED1在不同类群中分化明显,但其蛋白结构与功能依然高度保守,推测在被子植物的生殖进化中发挥核心作用。这种跨谱系的结构与功能保守性表明,DDR与SMPED1在被子植物基因组演化及性状分化中具有重要的进化意义。

图4:生姜及相关家族DDR的起源
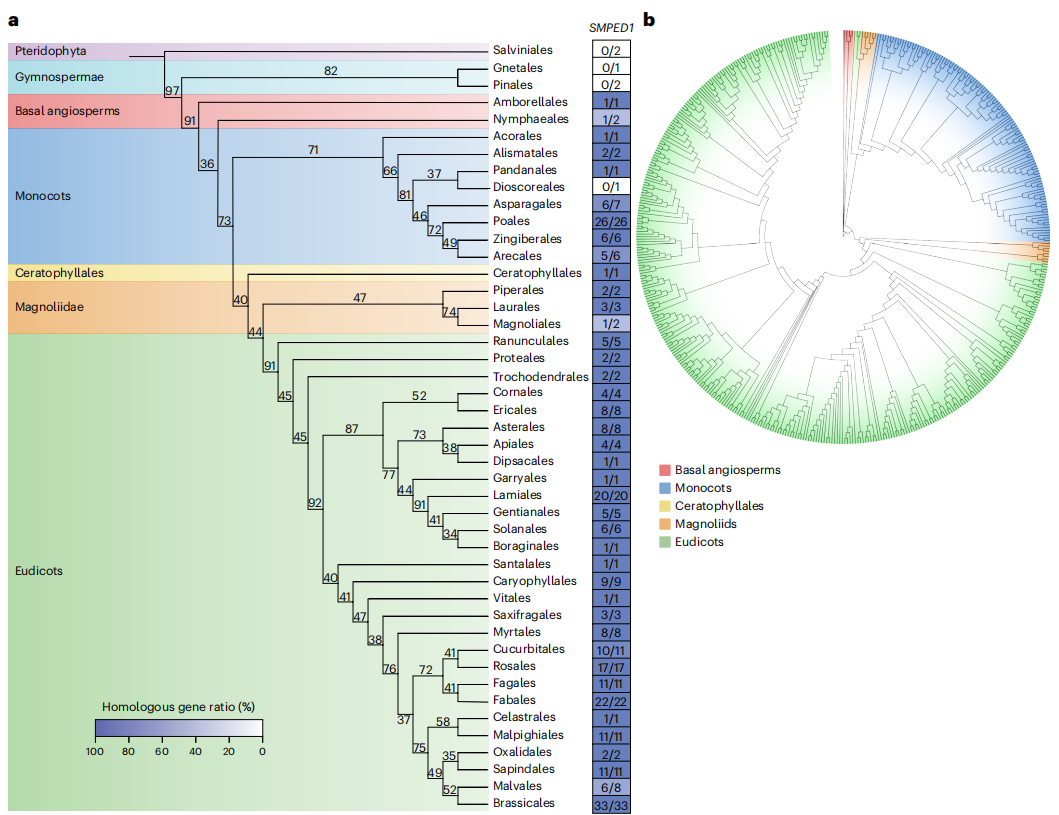
图5:被子植物中推测的DDR的进化
结语
本研究从基因层面阐明了姜科植物马来良姜的雌性异熟现象的遗传基础。通过对该物种单倍型基因组构建及GWAS分析,锁定了一个名为SMPED1的基因。转录组分析以及过表达和下调表达株系的结果表明,SMPED1基因可通过其在花药和柱头中的节律性表达,调节植物的性别功能。这一基因的功能在被子植物中广泛保守,表明其在植物繁殖中的广泛作用。研究还探讨SMPED1的进化,发现该基因在不同的被子植物群体中存在,揭示了其在植物性别系统中的重要性,为我们理解植物性别多态性的遗传机制做出了重要贡献。
华命生物产品服务一览

华命生物目前已开通微信公众号、抖音、知乎、B站、小红书等线上平台,欢迎感兴趣的老师扫码关注了解更多内容!
