

Plant J精选 | 大叶杨单倍型T2T基因组揭示着丝粒演化机制
着丝粒是染色体分离的重要结构,其序列和组成在不同植物中差异显著。除卫星重复序列外,转座元件(TEs)在着丝粒结构中也发挥关键作用。虽然模式植物研究较多,但多年生树种如杨树的着丝进化机制仍不清楚。随着长读长测序和 Hi-C 技术的发展,端粒到端粒(T2T)组装为解析植物着丝粒结构与演化提供了新契机。
南京林业大学的科研团队于2025年9月29日在期刊The Plant Journal上发表了一篇题目为“Haplotype-resolved telomere-to-telomere genome assembly of Populus lasiocarpa unveils retrotransposon-driven centromere evolution”的研究论文,组装出大叶杨高质量T2T级基因组组装,并进一步研究着丝粒的结构和演化,为植物基因组进化提供了新的见解。

一、PLAS 着丝粒的完整组装
本研究针对大叶杨Populus lasiocarpa进行T2T高质量基因组组装及着丝粒精确解析。通过整合PacBio HiFi、ONT超长读段、Illumina短读段和Hi-C数据,获得了单倍型分型的二倍体基因组,总长约881.7 Mb,包含38条染色体,其中36条实现端粒到端粒组装。基于CENH3 ChIP-Seq数据共鉴定出38个功能性着丝粒,除Chr08和Chr14因高密度45S rDNA阵列导致短臂不完整外,其余均完整。两单倍型的N50分别为23.3 Mb和24.5 Mb,BUSCO完整性超过99%,表明组装质量卓越。
甲基化与重复序列分析表明, TEs占基因组一半以上,着丝粒区域显著富集RLG/RIL类反转录转座子,并在CG、CHG和CHH位点表现出高甲基化状态。与模式植物富卫星DNA的着丝粒不同,大叶杨着丝粒主要由转座序列构成。基因注释共识别约3.6万个蛋白编码基因,BUSCO完整性达97.8%以上,其中数百个基因在着丝粒区具有较高表达活性。
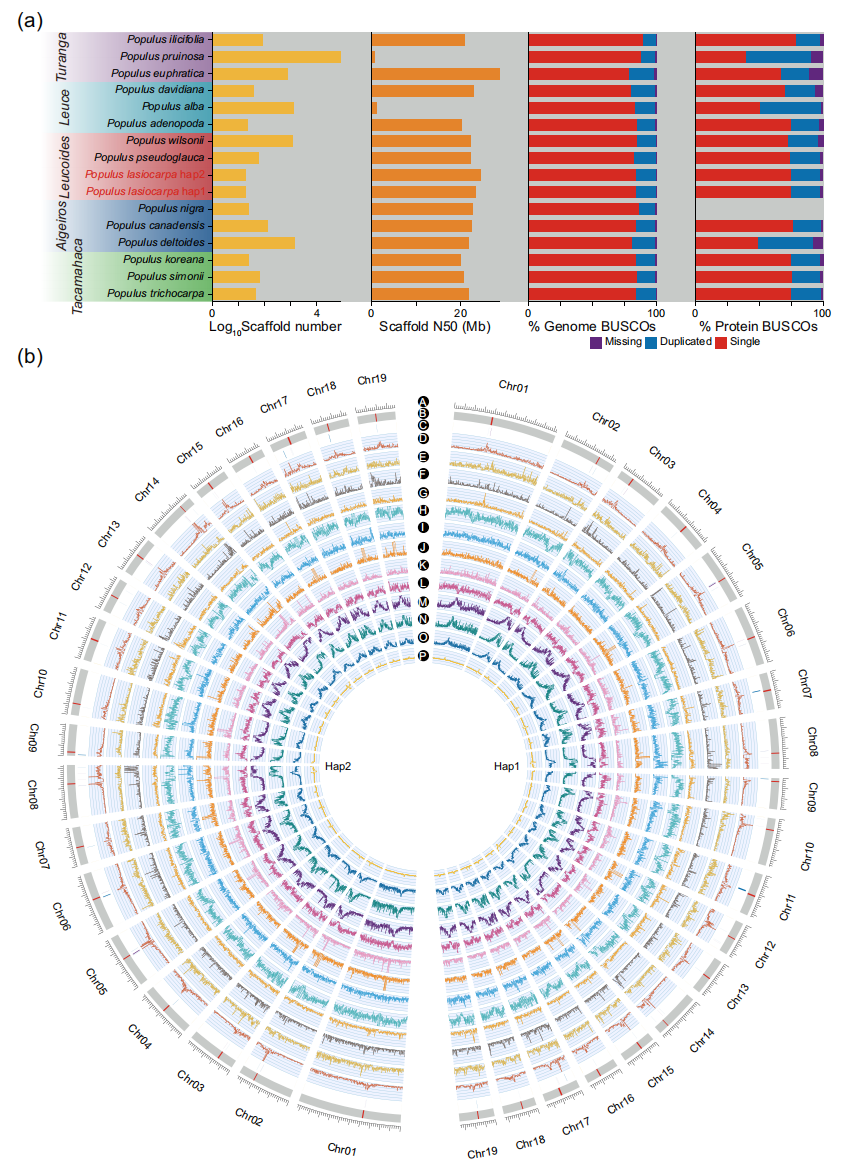
图1:PLAS单倍型基因组的组装和注释
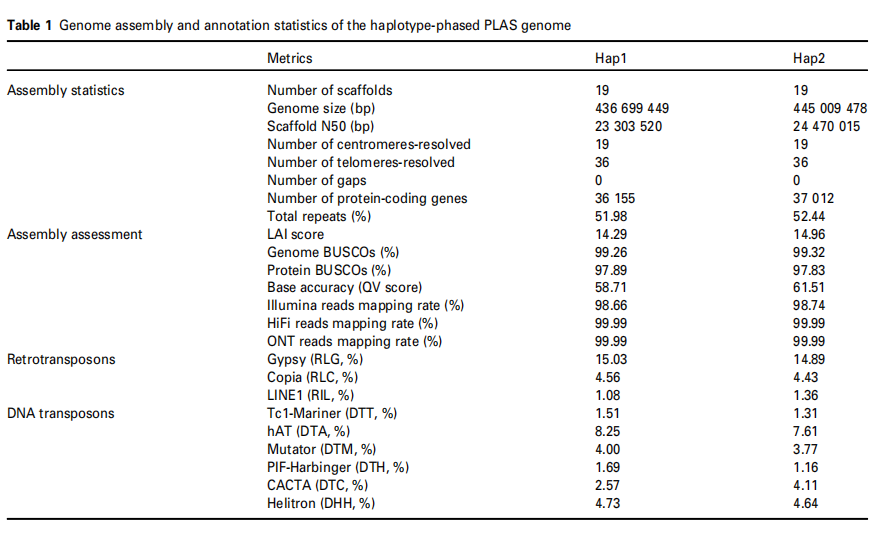
表1:单倍型阶段PLAS基因组的基因组组装和注释统计
二、PLAS着丝粒的定位和大小
本研究在大叶杨中实现了全部38个功能性着丝粒的精确定位,确认其为典型的单着丝粒型植物。所有着丝粒均被ONT超长读段完整覆盖,在hap1和hap2中的平均长度分别为553 kb和544 kb。根据染色体臂比,大叶杨染色体可分为端着丝粒型、近端着丝粒型、中近着丝粒型和中着丝粒型,同源染色体对在两单倍型间具有高度一致的臂比值,表现出出高度保守的结构特征。着丝粒内部由交错分布的CENH3与H3富集区构成,呈镶嵌状结构,体现出反转录转座子富集型着丝粒的保守组织模式。此外,卫星与微卫星重复序列在功能性着丝粒中几乎缺失,而主要分布于近着丝粒区。在功能性着丝粒中共识别出749个蛋白编码基因,其中约115个在至少一种组织中具有活跃表达,主要参与光合作用、脂肪酸合成和过氧化物酶体功能,功能性着丝粒基因的DNA甲基化水平显著高于常染色体臂。
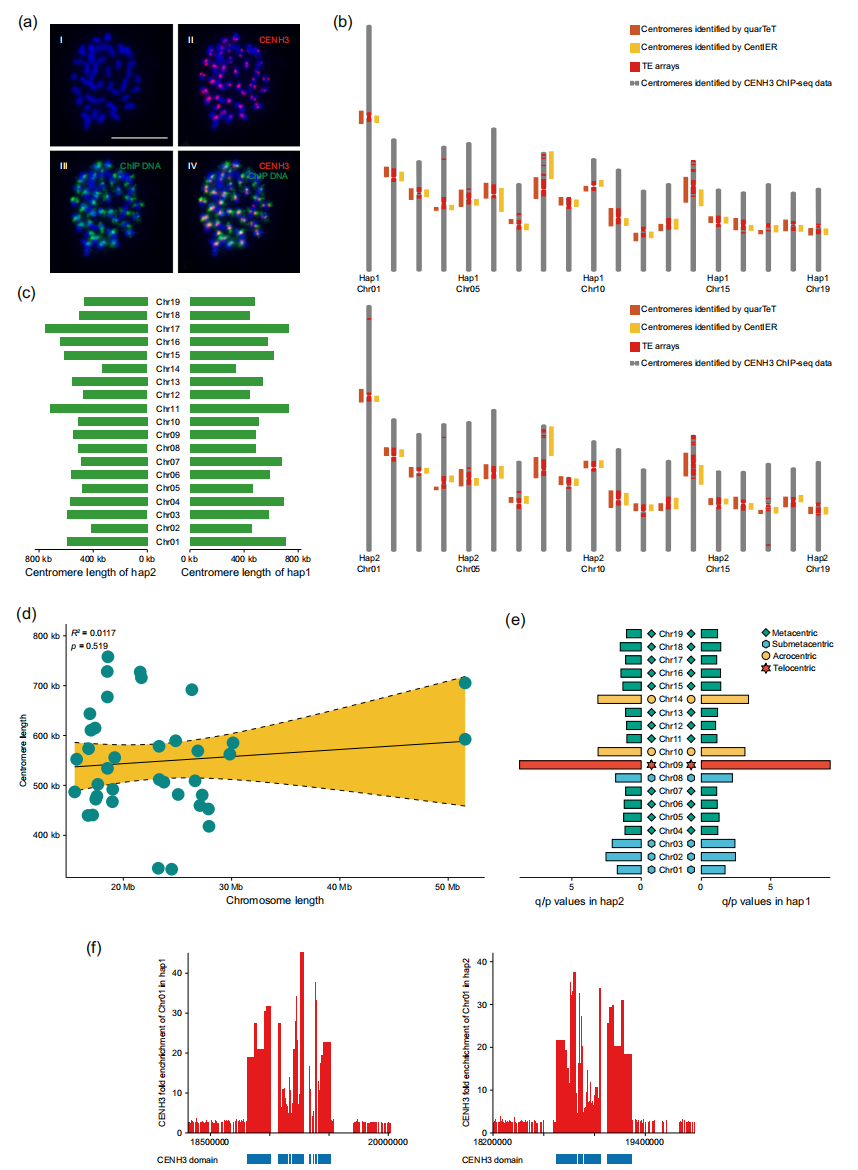
图2:PLAS着丝粒的定位
三、PLAS着丝粒表观遗传特征
作者进一步探究了着丝粒的表观遗传特征,结果显示着丝粒区域相较染色体臂具有显著的DNA高甲基化,是跨物种保守的特征。在全部38条染色体的H3染色质区中,RLG与RIL反转录转座子形成嵌套TE阵列,而这些区域缺乏CENH3核小体结合。嵌套TE的侵入会降低DNA甲基化水平,破坏原有甲基化模式。研究表明,CENH3结合主要受表观遗传状态而非序列控制,且其信号与甲基化水平相关性较弱,说明组蛋白修饰等其他表观机制亦在CENH3结合中发挥重要作用。
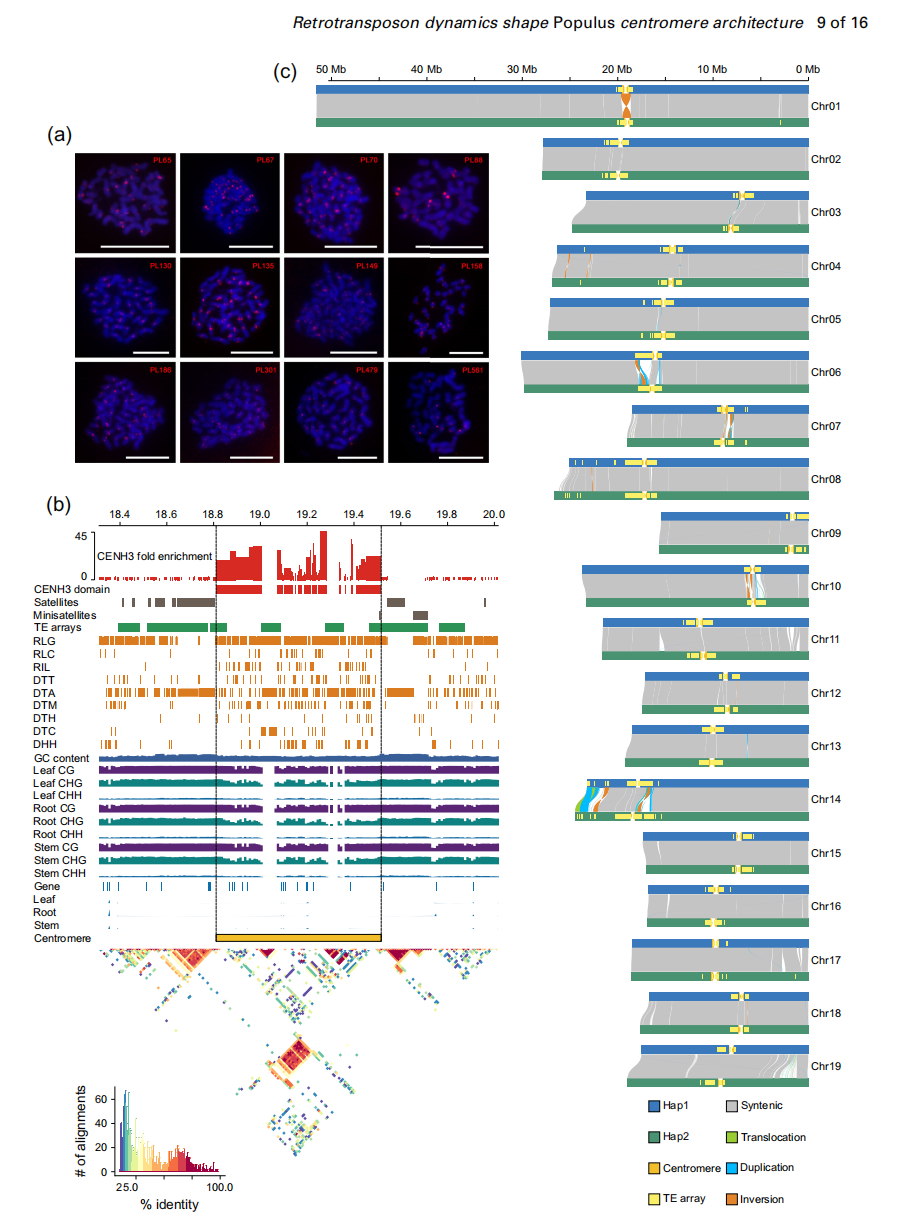
图3:PLA着丝粒的结构结构
四、杨属着丝粒的比较分析
作者进一步结合已发表的毛果杨Populus trichocarpa的T2T单倍型组装,对杨属植物着丝粒进行了高精度比较分析。结果显示,毛果杨与PLAS在着丝粒大小、染色质结构(交错分布的CENH3/H3区域)及转座元件组成(以RLG/RIL反转录转座子为主,缺乏卫星重复序列)方面高度相似,表明二者在表观遗传和功能层面具有显著保守性。尽管整体结构相似,但两者的着丝粒DNA序列存在显著分化,不同染色体间的相似性差异与嵌套型RLG/RIL转座子阵列密切相关。嵌套转座子的插入破坏序列完整性并促进着丝粒进化,而非嵌套型转座子无此不稳定效应。作者指出,嵌套反转录转座子的组织结构是驱动杨属着丝粒演化的重要因素。
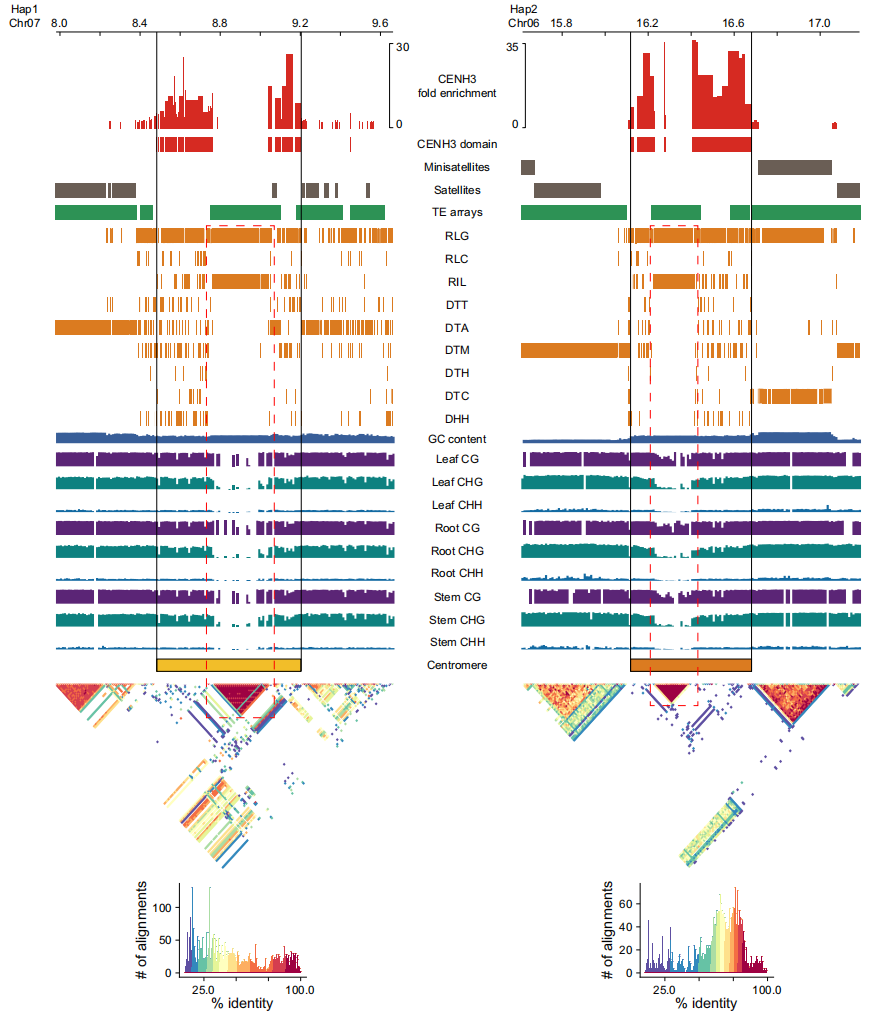
图4:PLAS着丝粒的表观遗传景观
五、杨属着丝粒进化模型
作者进一步提出了“反转录转座子驱动的杨属着丝粒进化模型”。经典观点认为,TE富集型着丝粒代表进化的早期阶段,而卫星重复富集型着丝粒则是进化成熟的形态。在拟南芥中,KARMA机制通过清除入侵的ATHILA转座子,促进高阶卫星重复的扩增,从而形成稳定的着丝粒结构。与此相比,杨属着丝粒以RLG/RIL类反转录转座子为主,体现了从TE富集向TR主导过渡的早期阶段。该模型认为,自主型TE的持续转座扰乱DNA甲基化与组蛋白修饰,削弱CENH3的结合能力,进而诱导嵌套TE阵列的扩张;这一过程可能导致原有着丝粒失活并激活潜在位点形成新生着丝粒,而富含TE/TR且甲基化水平较高的近着丝粒区域成为新生着丝粒形成的“热点区域”。TR富集型着丝粒可通过KARMA机制持续清除TE,逐步演化为稳定的单中心卫星结构;而TE富集型着丝粒则经历反复的“失活-重建”循环,最终在TR富集区形成新的中心。该模型揭示了着丝粒功能稳定而序列快速更替的进化悖论,并指出反转录转座子与卫星重复之间长期“拉锯式”动态平衡在染色体结构进化与核型多样性形成中的关键作用。
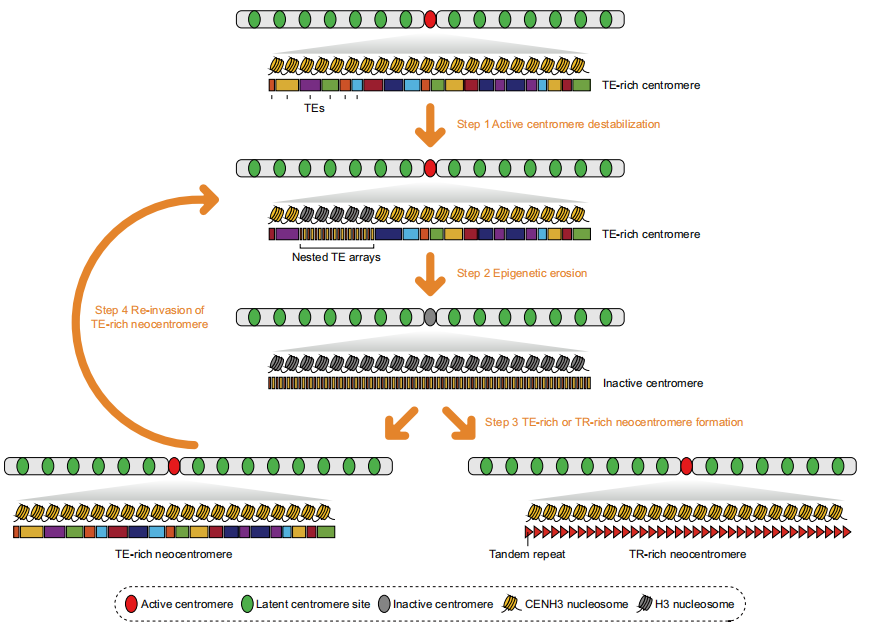
图5:反转录转座子驱动着丝粒进化模型
结语
本研究完成了杨树T2T单倍型基因组的组装,重点分析了其着丝粒区的进化机制。作者发现,杨树的着丝粒并不像其他模式植物那样富含卫星DNA,而是由反转录转座子主导,尤其是富含RLG和RIL,它们在功能性着丝粒区域内形成复杂的嵌套转座子阵列,破坏了其结构完整性并驱动了其进化。作者提出了一种反转录转座子驱动的着丝粒进化模型,认为自主反转录转座子的不断转座,通过表观遗传侵蚀使功能性着丝粒不稳定,从而触发新着丝粒的形成。这些新着丝粒通常在富含转座子和串联重复的近着丝粒区域形成,并且可能随着反转录转座子的再度入侵而不断发生变动。本研究提出了一个着丝粒模式新的视角,强调了反转录转座子在植物基因组演化中的关键作用,促进了对杨树以及其他植物着丝粒多样性的深刻理解。
华命生物产品服务一览

华命生物目前已开通微信公众号、抖音、知乎、B站、小红书等线上平台,欢迎感兴趣的老师扫码关注了解更多内容!
