

PBJ精选 | 水稻泛基因组解码籼粳杂种优势
杂种优势是指F₁杂交种因为亲本基因互补而表现更好,这一现象对作物育种很重要,但它到底由哪些遗传差异造成,长期以来并不完全清楚。早期研究主要关注SNP、InDel等小范围变异,发现亲本差异和杂交优势往往相关。近年随着高质量基因组测序与组装发展,研究者开始注意到更大尺度的结构变异(SV)也可能深刻影响性状与杂种优势,但在水稻中其作用仍缺乏系统解析。
武汉大学生命科学学院黄文超研究团队于2025年11月22日在国际著名期刊Plant Biotechnology Journal上发表标题为“Structural Variations Contribute to Subspeciation and Yield Heterosis in Rice”的研究论文,系统构建水稻杂交泛基因组和结构变异图谱,证明结构变异通过重塑基因表达决定籼粳分化、杂种优势和生殖隔离,并锁定一批可用于优化籼粳亲本设计的关键位点。

一、12 个代表性水稻自交系的基因组组装与注释
本研究从223个水稻自交系中选取12个具有代表性的indica、中间型和japonica基因型,用于解析杂种优势的遗传基础。这些材料具有显著的表型差异,来源覆盖中国、印度和意大利。基于全互交设计构建的120个F₁杂交种显示种间杂交(indica × japonica)普遍表现为结实率降低、株高升高,杂种优势水平也与种内杂交显著不同。利用PacBio和Hi-C技术,作者对12个IL品种进行了高质量基因组组装,所得基因组的contigN50、锚定率、BUSCO完整度等指标均表明其质量优异,重复序列占基因组的55%以上,YB和Balilla分别注释到约4.4万与4.37万基因。进一步对Phr1基因的分析显示,其18/29bp缺失与酚反应表型密切相关:缺失突变的材料在酚处理后迅速变黑,而其他材料在124天后仍保持浅色,验证了新组装基因组的准确性,并强调了Phr1关键变异在表型中的作用。
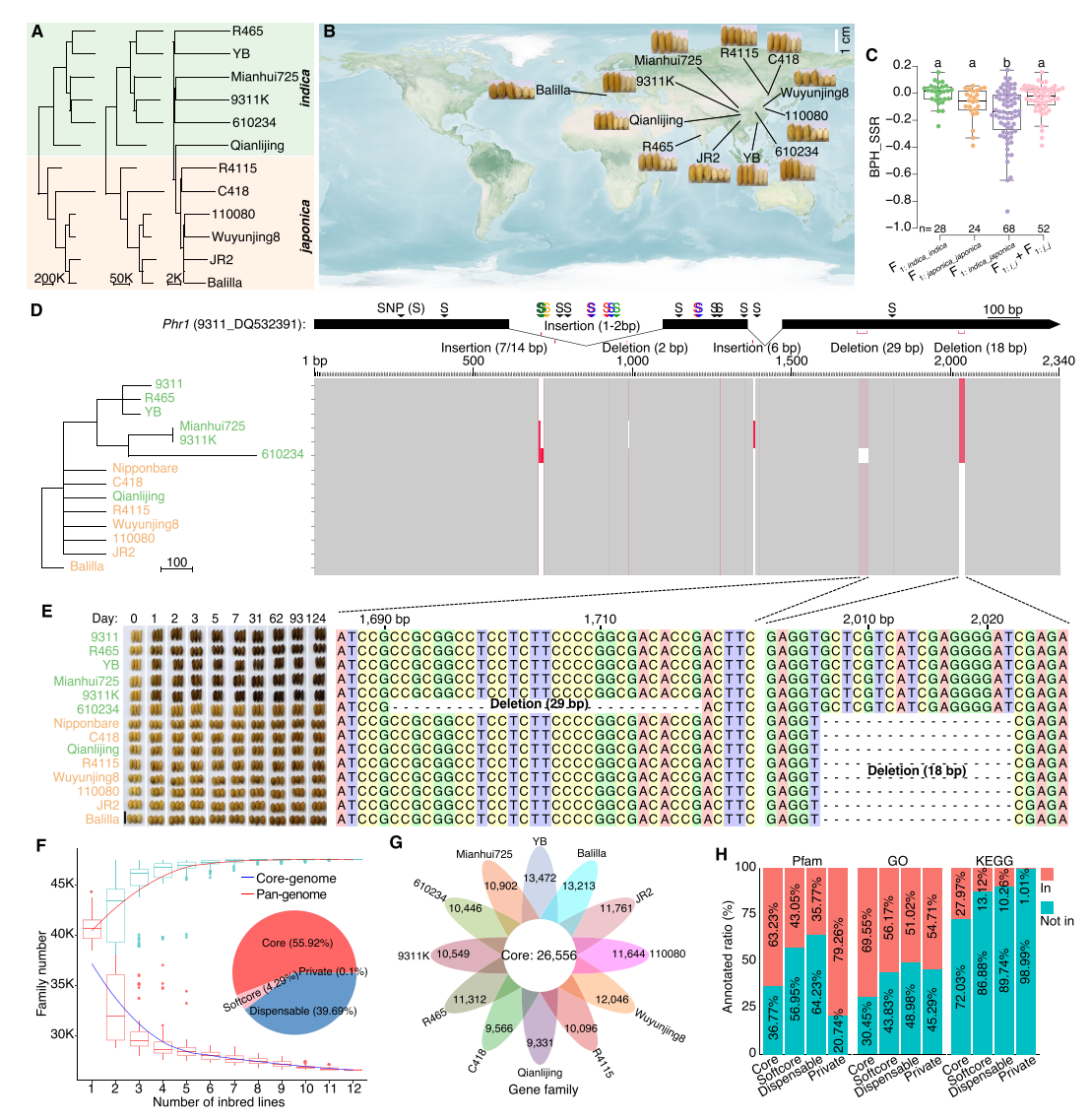
图1: 12个水稻自交系的基因组组装及泛基因组
二、12 个自交系的泛基因组构建与泛结构变异分析
本研究利用12个水稻自交系构建了泛基因组,共识别47,490个基因家族,其中核心基因家族约占56%,可变家族约占40%,特有家族极少。基因家族数量在加入10个基因组后趋于稳定,说明材料具有代表性;而核心基因随样本增加而减少,反映出不同自交系的特异性。非核心基因家族在不同材料间差异显著,且核心基因在Pfam、GO、KEGG的注释比例最高,说明其功能高度保守。
将参考基因组YB与其他自交系(ILs)比较后,检测到大量结构变异,包括存在/缺失、拷贝数变异、易位及倒位。SV的数量与长度均呈indica向japonica增加趋势,japonica含量显著更高。6号、8号染色体上发现的大型倒位其可能推动亚种分化。与泛基因组不同,泛SV在11个样本时仍未达平台期,显示其高度多样性。多数SV富集于DNA转座子和LTR区域,可能调控基因组非编码区功能。
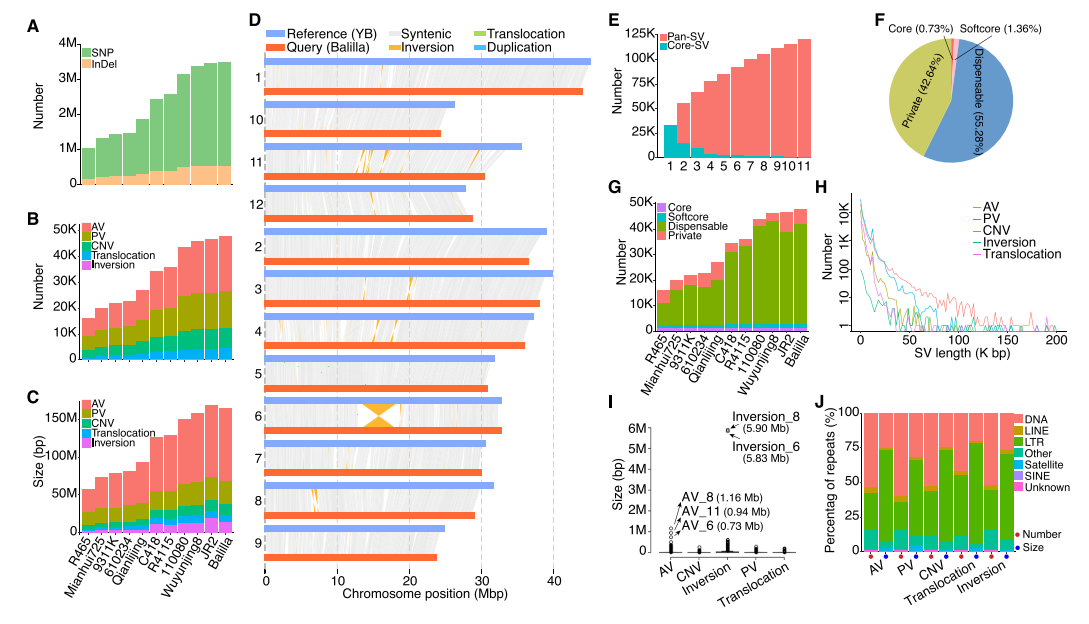
图2:12条近交系的基因组变异
三、结构变异与 17 项农艺性状杂种优势的关联
本研究比较了12个自交系之间的基因组差异,找出了它们各自独有或共享的变异。作者发现,任意两个自交系之间平均有约201万SNP、39万InDel和4万SV。特有变异数量彼此相关,多位于基因间区;倒位中有近78%出现在外显子。不同变异类别构建的系统关系不一致,说明它们在基因组分化中发挥着不同作用。作者进一步将亲本变异与17个性状的杂种优势进行比较,发现特有变异更能影响杂交后的性状表现,而SNP和InDel之间的差异不大。虽然特有SV数量少,但对杂种优势的作用与SNP/InDel相当,可能因影响范围更广。倒位数量虽最少,却与株高杂种优势关系最强。特有变异与结实率杂种优势呈负相关,说明存在生殖隔离;其中特有SV增加会明显降低籼×粳F₁的结实率。
本研究还发现,籼粳杂交亲本含有更多特有SV,这些变异与分蘖数杂种优势更相关,可能在种间杂种优势和增产潜力中的重要作用。但它们与产量杂种优势的关系较弱,可能是因为结实率下降所致。
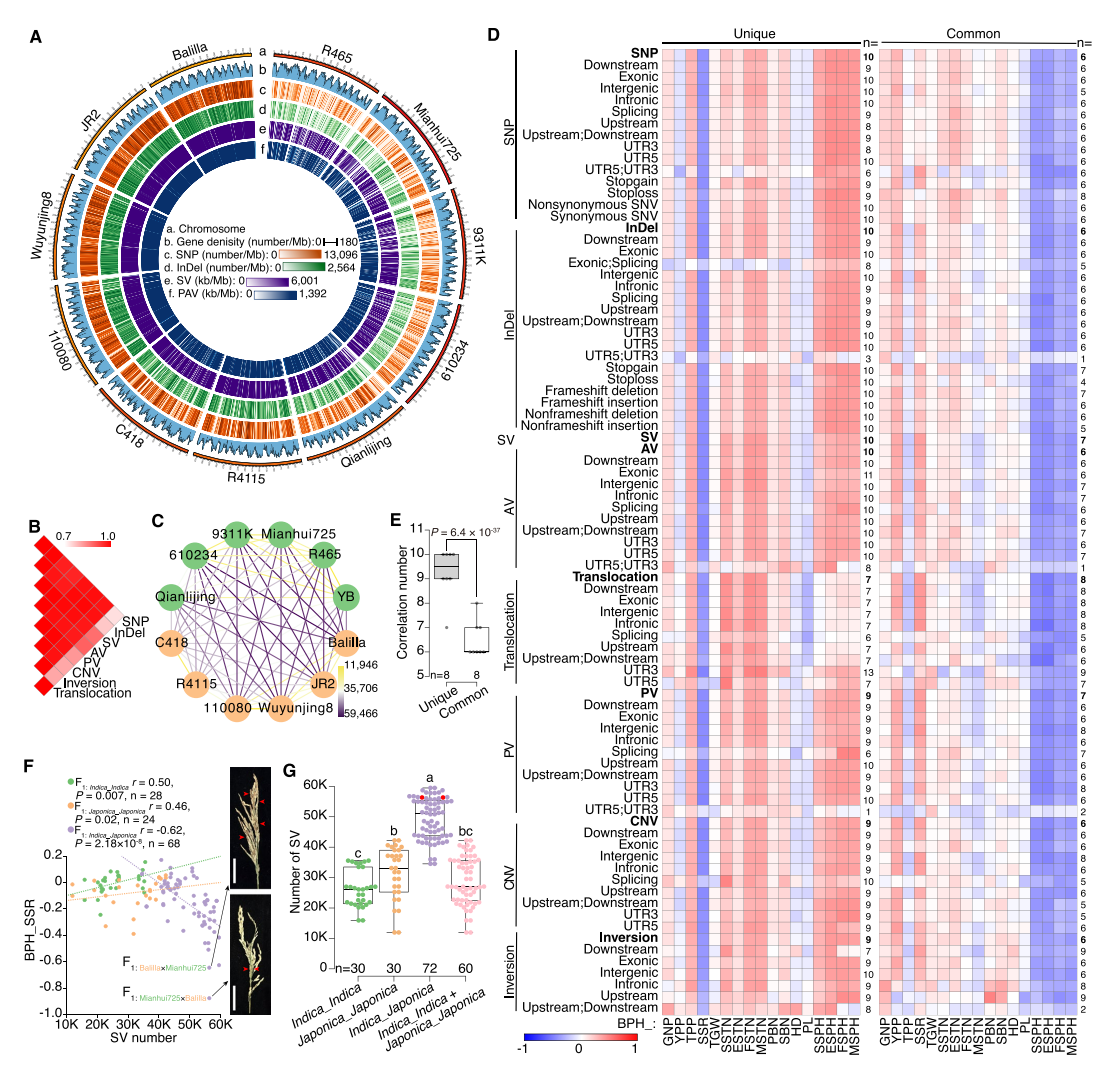
图3:父母基因组变异与17种农艺性状杂种优势的相关性
四、S5-ORF5 缺失变异驱动结实率杂种优势
作者针对籼粳杂交不亲和关键位点S5进行了系统分析。在对S5-ORF3、S5-ORF4、S5-ORF5的序列比较中发现,YB与黔礼粳在系统发育上聚为一簇,并携带S5-ORF5中的94bp缺失变异,同时伴随10bp与42bp缺失。这些变异紧密连锁,仅存在于两者中,被认为能移除杀伤效应,使籼粳F₁胚囊得以存活,因此被鉴定为广亲和品种(WCVs)。功能验证表明一般籼粳F₁结实率显著低于籼籼与粳粳,但当YB或黔礼粳作为亲本时,其籼粳F₁结实率与籼籼F₁相当,杂种优势亦同步提升。由此证实YB与黔礼粳为籼型广亲和品种,且94bpAV是提升籼粳结实率与杂种优势的关键因素。
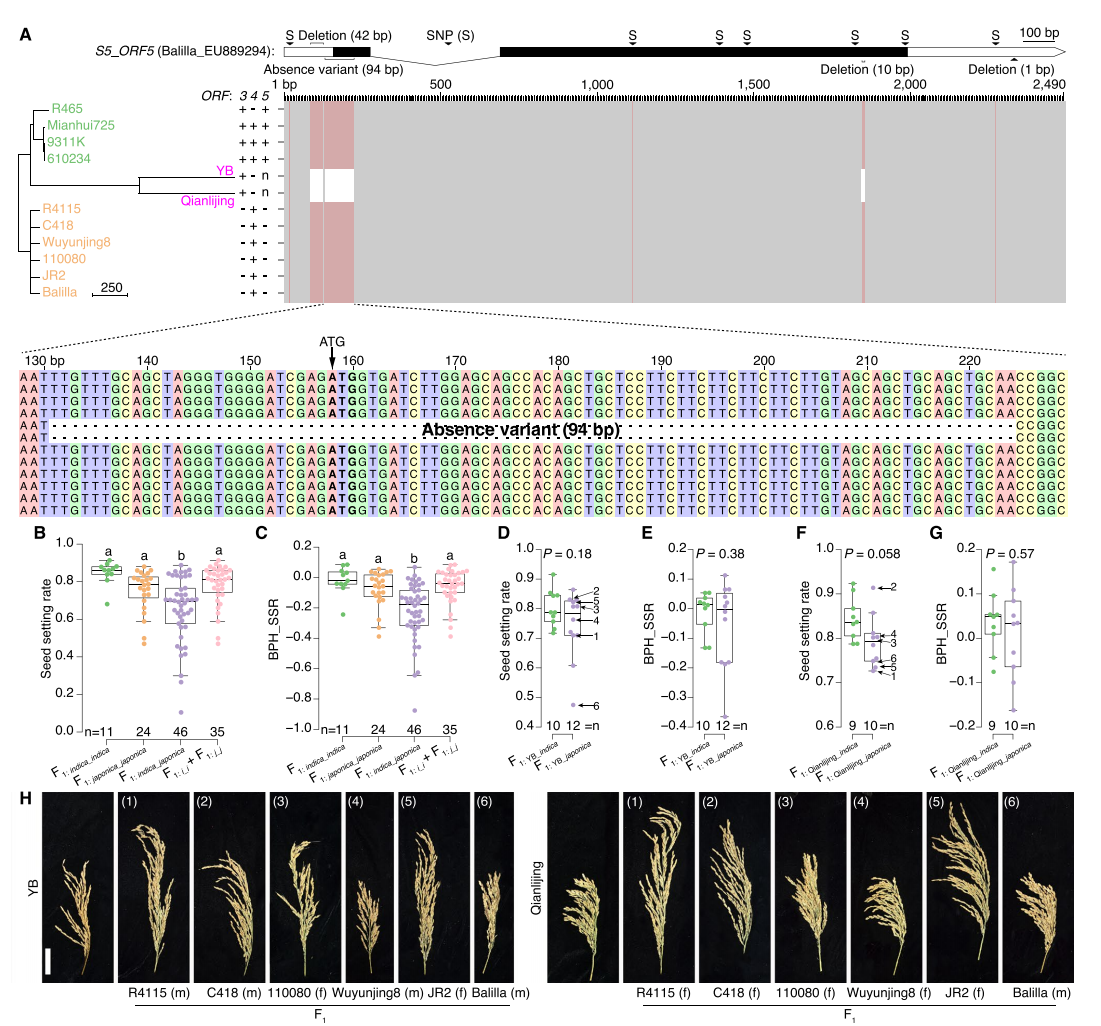
图4:S5-ORF5中AVs对籽粒结实率杂种优势的贡献
五、OsMADS1 验证及 OsBZR1 变异在杂种优势中的作用
研究鉴定出OsMADS1的三个单倍型,并检测到多种SNP和结构变异。进一步分析表明,由不同单倍型构成的杂合F1在产量上显著优于同型组合,说明OsMADS1的杂合性对杂种优势形成具有重要作用。作者同时关注到OsBZR1,这一油菜素内酯信号通路的关键因子参与了籼稻和粳稻的表型分化,其启动子中存在重要的4808bp反转座子插入。系统发育分析将OsBZR1分为籼型、粳型和JR2型,并检测到82个SNP、266bp的粳稻特有变异以及4808bp的籼稻特有变异等多种类型。表达分析显示,粳稻的OsBZR1表达显著高于籼稻,并且亲本表达量越低的组合,其F1杂种优势越强。
在田间性状分析中,作者发现籼型与粳型之间的杂交差异不明显,而籼型与JR2型(i/JR2)的F1在产量、株高、分蘖数等多项指标上表现出最强的杂种优势。相较之下,粳型与JR2型的组合并未出现明显优势。综上,本研究认为i/JR2的杂合是形成高产杂种优势的关键因素。
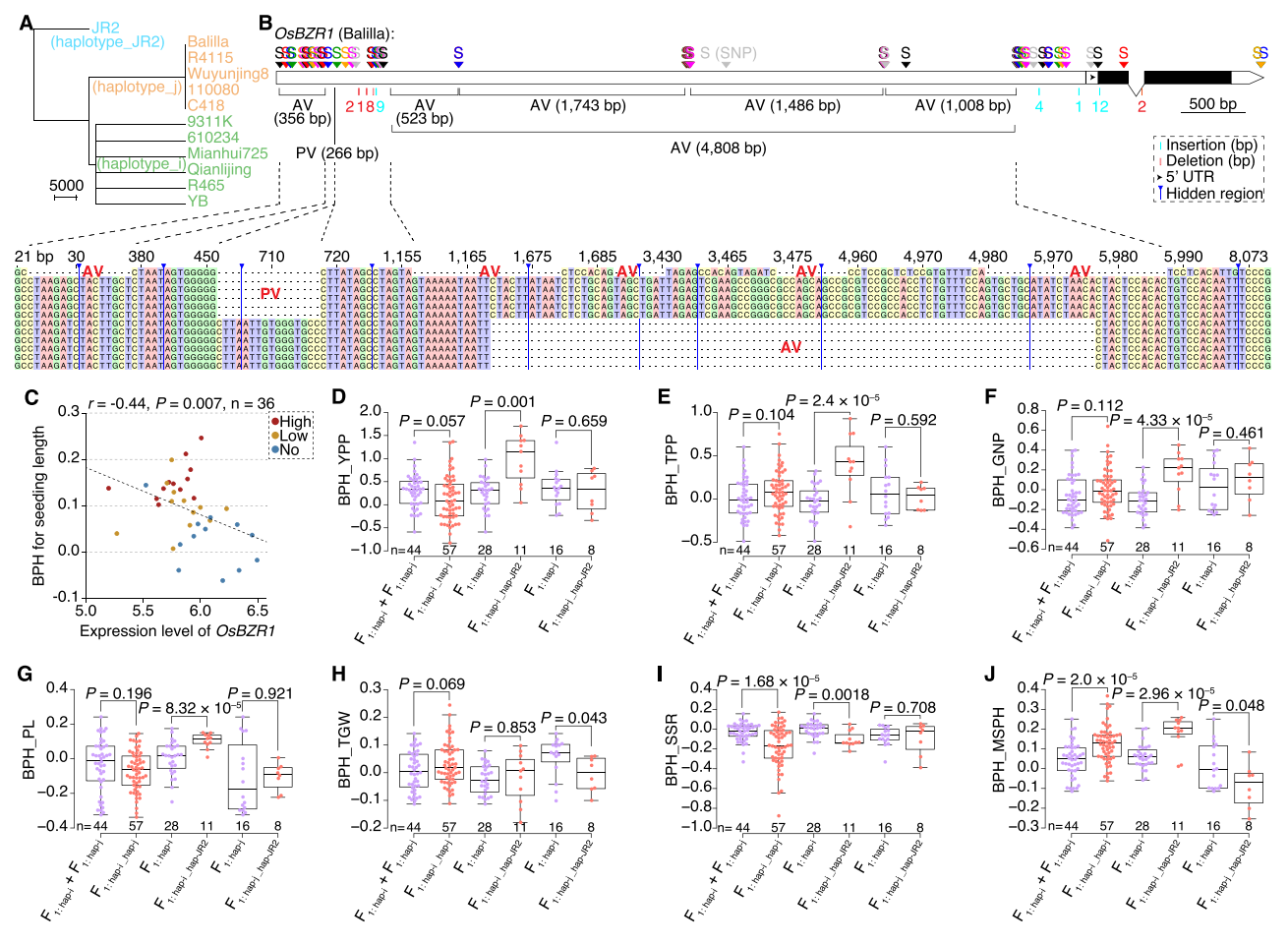
图5:OsBZR1中SVs对产量杂种优势的贡献
六、结构变异通过调控基因表达解释杂种优势差异
作者通过对12个自交系幼苗的转录组分析,系统揭示了结构变异如何影响基因表达并驱动杂种优势形成。差异表达分析显示,这些材料可被分为两个主要类群,说明基因组层面的差异已扩展到表达层面。进一步聚焦大型倒位与缺失区域,作者发现inversion_6能清晰区分籼、粳类型,这里的基因呈现的加性与部分显性效应能够用来预测杂种优势;而inversion_8虽然涉及的差异基因不多,但其表达模式仍然具有很强的解释能力。在三类大型缺失中,AV_6的预测表现甚至可以和使用整个基因组的差异表达基因或倒位区域的结果相媲美。
为了评估亲本间结构变异对杂交后F₁表达的影响,作者将1759个差异表达基因与不同变异类型的数量一起进行PLS分析,结果表明结构变异的预测能力明显强于SNP和InDel,其中尤以易位效应最为突出。不同易位事件在不同遗传背景中会导致完全相反的表达分组,说明其效应具有背景依赖性。进一步分析表明,外显子缺失、5'UTR拷贝数变化、剪接位点变异以及倒位区域的基因间区,都能更有效地预测杂种优势。对典型结构变异的单倍型进一步分析后发现,拥有杂合单倍型的材料往往表现出更强的杂种优势,支持水稻中“超显性模型”的存在。
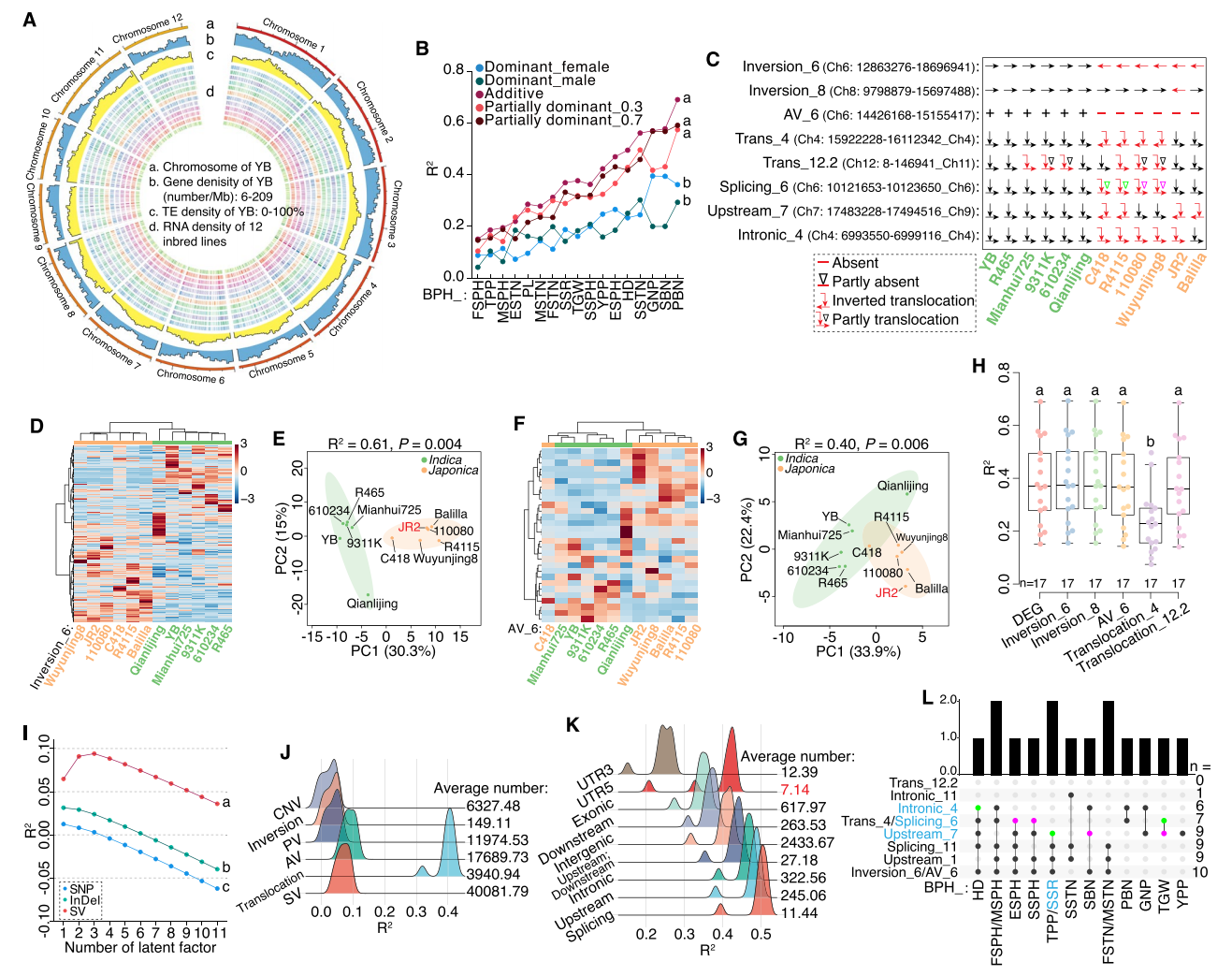
图6:基因组结构变异、基因表达与杂种优势的关系
结语
本研究组装了12份代表性籼稻、粳稻及中间类型材料的高质量基因组,构建了水稻泛基因组和结构变异图谱,证明插入缺失、倒位和易位等结构变异是籼粳分化和重要农艺性状差异的关键基础。研究团队又构建120个F1杂交组合,测定17个农艺性状,发现亲本之间结构变异越多,杂种优势整体越明显。而籼粳间杂交如果变异过多,结实率会明显下降,说明存在生殖隔离。进一步解析到几个关键位点,例如S5-ORF5的缺失等广亲和型等位变异可以缓解籼粳不亲和、提升结实率,而OsMADS1和OsBZR1的不同单倍型组合与产量和株高等杂种优势紧密相关。转录组分析显示,大尺度的结构变异能够重塑附近基因表达,杂合状态普遍优于纯合状态,支持以结构变异杂合度为核心的超显性模型,为设计更优的籼粳亲本组合和利用杂种优势提供了理论基础。